Verschlüsselter Code: Übererregbarkeit des Trigeminussystems
Verschlüsselter Code: Übererregbarkeit des Trigeminussystems
Zusammenfassung

Der 32-jährige Patient, der unter ausgeprägtem nächtlichem und täglichem Bruxismus sowie orofazialem Schmerz (OP) vorwiegend in den temporoparietalen Regionen mit einer höheren Intensität und Häufigkeit auf der linken Seite des Gesichts leidet, unterzog sich dem diagnostischen Modell Masticationpedia, das die Maschinensprache des Zentralnervensystems im Rahmen der "Hyperexitabilität" des Zentralnervensystems mit besonderem Bezug auf den mittleren Hirnstammtrigeminusbereich entschlüsselte. Diese "Hyperexitabilität" wurde durch eine elektrophysiologische Methode namens "Erholungszyklus des inhibitorischen Masseterreflexes" verifiziert, die eine übertriebene Erholung der Ruhephase hervorrief, die durch den zweiten elektrischen Reiz namens "Stimulus-Test" ausgelöst wurde. Dieser neurophysiologische Zustand veranlasste den Arzt, eine MRT des Gehirns zu beantragen, die eine "Pinealis Cavernosa" zeigt. Zusammenfassend ist "Bruxismus" eine Form der funktionellen Instabilität der neuronalen Erregbarkeit und/oder organisch, daher nicht ausschließlich auf die Zahnmedizin bezogen. Die Fortsetzung des Phänomens, seine Intensivierung und die Hartnäckigkeit, es mit zahnärztlichen Behandlungen zu behandeln, ohne den "Zustand" des Systems genauer zu untersuchen, könnte ernsthaft sein und eine ungünstige Prognose haben.
Einleitung
Wir haben daher den Abschnitt des Cognitive Neural Network' erreicht, das als 'RNC' abgekürzt wird und für die Diagnose des Falls unserer 'Mary Poppins' im Kapitel 'Verschlüsselter Code: Ephaptische Übertragung' vorgestellt wurde. Wir werden es erneut als Diagnosemodell vorschlagen, um den Leser mit dem Verfahren vertraut zu machen, das einfach, intuitiv, aber unerlässlich bei klinischen Fällen mit komplexer Diagnose wie unserem Patienten 'Bruxer' ist. Unser Ausgangspunkt ist daher der Endpunkt der Phase vor dem 'RNC', nämlich die diskriminierende Phase der Kontexte ( Kohärenz-Demarker). Das geringe diagnostische Gewicht, das sich aus den neurologischen Aussagen ergibt, bezieht sich tatsächlich nur auf einen bescheidenen Unterschied in der Amplitude des Kieferreflexes. Auch in diesem Fall kann uns das Cognitive Neural Network (CNN) dabei helfen, den Maschinensprachencode zu fokussieren und zu entschlüsseln. Wir folgen daher demselben Verfahren, das bereits ausführlich im Kapitel 'Verschlüsselter Code: Ephaptische Übertragung' beschrieben wurde, und erhalten das folgende Ergebnis:


Bruxism (4398), trigeminal system (29), abnormality ( 5), excitability ( 3) The excitability of the trigeminal motor system in sleep bruxism: a transcranial magnetic stimulation and brainstem reflex study


Diagnostische Sequenzen
1 Schritt: CNN-Sequenz
- -Kohärenz-Demarker: Wie bereits beschrieben, besteht der erste Schritt aus einem Initialisierungsbefehl der Netzwerkanalyse, der sich tatsächlich aus einer vorherigen kognitiven Verarbeitung der Aussagen im zahnärztlichen Kontext und im neurologischen Kontext γn ableitet, denen der ' -Kohärenz-Demarker' ein absolutes Gewicht verliehen hat, wodurch der zahnärztliche Kontext effektiv aus dem Prozess eliminiert wird. Aus den neurologischen Aussagen ergibt sich, dass der 'Zustand' des Trigeminusnervensystems in Bezug auf die Amplitude des Kieferreflexes relativ asymmetrisch ist, wobei ein Durchschnitt von vorliegt. Dies erlaubt es nicht, den anfänglichen rein neurologischen Befehl in die Pubmed-Datenbank einzugeben, wie es für den vorherigen klinischen Fall von Mary Poppins durchgeführt wurde. Der Initialisierungsbefehl wird daher 'Bruxismus' sein, der sich auf beide Datensätze (zahnärztlich und neurologisch) bezieht.


- 1st Schleife geöffnet: Dieser "Initialisierungsbefehl" wird daher als anfängliche Eingabe für die Pubmed-Datenbank betrachtet, die mit 4398 klinischen und experimentellen Daten antwortet, die dem Kliniker zur Verfügung stehen. Die Eröffnung der ersten tatsächlichen kognitiven Analyse erfolgt präzise auf der Analyse des ersten Ergebnisses des 'CNN', das 'Bruxismus' entspricht. In dieser Phase, angesichts der Negativität des zahnärztlichen Berichts und der minimalen Positivität des neurologischen Kontexts, wäre es notwendig, eine neurologische Komponente zu identifizieren, für die das 'Das trigeminale System' als erster offener Regelkreis hinzugefügt wird.
- 2st Schleife geöffnet: Der Prozess wird fortgesetzt, indem der Fokus immer mehr auf die Schlüsselwörter gelegt wird, die mit unseren trigeminal-spezifischen Kontextergebnisdaten übereinstimmen, die wir mit einem Begriff 'Abnormalität' vervollständigen sollten. Dieser Begriff wird die 2. offene Schleife mit 5 spezifischen Elementen durchführen. An diesem Punkt sollte man kognitiv den Aufwand betreiben, alle 5 Artikel zu bewerten, um einige klinische oder laboratorische Hinweise extrapolieren zu können, die für die Entschlüsselung des maschinensprachlichen Codes des zentralen Nervensystems erforderlich sind. Die Bewertung der Artikel ergab ein Phänomen, das möglicherweise in einigen Fällen von Bruxismus vorhanden ist, nämlich eine veränderte Erregbarkeit des Trigeminussystems. Daher wurde der Begriff 'Erregbarkeit' in das Netzwerk in der 3. offenen Schleife eingefügt.
- 3st Schleife geöffnet:In dieser Phase lieferten die "Erregbarkeits"-Daten 3 sehr signifikante Artikel zurück, die darauf hinweisen, wie der Erregbarkeitsgrad des trigeminalen Zentralnervensystems durch eine elektrophysiologische Technik namens "Erholungszyklus" des masseterischen inhibitorischen Reflexes getestet werden kann und als rcMIR abgekürzt wird. Offensichtlich wurde im vorliegenden Fall die Schleife des Netzwerks im ersten Artikel geschlossen, in dem diese Methodik erwähnt wird (der letzte Artikel in chronologischer Reihenfolge war unser). Aus diesem Artikel "Die Erregbarkeit des trigeminalen Motorsystems beim Schlafbruxismus: Eine transkranielle Magnetstimulation- und Hirnstammreflexstudie" geht hervor, dass bei Patienten, die Anzeichen und Symptome von nächtlichem Bruxismus (SB) berichtet haben, eine abnorme Erregbarkeit der trigeminalen motorischen Bahnen besteht. Diese erhöhte Erregbarkeit könnte auf eine veränderte Modulation inhibitorischer Hirnstammkreise und nicht auf veränderte kortikale Mechanismen zurückzuführen sein. Die Ergebnisse unterstützen die Idee, dass Bruxismus hauptsächlich zentral vermittelt wird und subkortikale Strukturen einbezieht.
2 Schritt: Erholungszyklus des inhibitorischen Masseter-Reflexes
Der Erholungszyklus des inhibitorischen Masseterreflexes (rcMIR) wurde durch die Erzeugung von Paaren von Reizen mit identischen Eigenschaften untersucht, die perkutan mit einem bipolaren elektrischen Stimulator geliefert wurden, der im Bereich des mentalen Nervs des Patienten positioniert war. Die Stimulation erfolgte mit Rechteckimpulsen, die in der Lage waren, einen klar definierten inhibitorischen Reflex zu erzeugen, der aus zwei stillen Perioden (SP) bestand, genannt SP1 und SP2, die durch ein Intervall der Erholung der EMG-"Interposed Activity" (IA) getrennt waren. Der erste Reiz (S1) wurde als Konditionierungsreiz betrachtet und der zweite (S2) als Testreiz. Das Intervall zwischen S1 und S2 wurde auf 150 ms festgelegt.
Der Proband wurde gebeten, die Zähne zusammenzubeißen, um maximale EMG-Aktivität zu erzeugen, und die Kontraktion mindestens 3 Sekunden lang aufrechtzuerhalten, mit visuellem und akustischem Feedback. Nach 60 Sekunden Ruhe wiederholte der Proband die Kontraktion 10 Mal. Das EMG-Signal wurde im direkt gleichgerichteten und gemittelten Modus aufgezeichnet. Die Platzierung der Ableitungselektroden war dieselbe wie die zur Aufzeichnung des Kieferreflexes verwendete, und die Parameter des Vorverstärkers wurden auf eine Zeitfensterbreite von 500 ms, 200 mV pro Teilung und eine Filterbandbreite von 50-1 kHz eingestellt. Die Latenzen und Dauern der SPs und der IA (Abbildung 1) wurden wie folgt berechnet:
- Um die Untersuchung zu vereinfachen, wurde der rcMIR durch elektrische Stimulation nur auf der linken Seite ausgelöst. Die EMG-Reaktionen entsprechen den EMG-Aufzeichnungen des rechten Masseters (Ch1) und des linken Masseters (Ch2). Auf den Spuren zeigt jeder Marker die Kanalnummer an, während die Buchstaben die Sequenzen von Latenzen anzeigen.
- Der Stimulus S1 teilt die Erfassung in Vor- und Nachanalyse auf und erzeugt die SPs und die IA.
- Der Stimulus S2, der 150 ms nach S1 geliefert wird, der als Interstimulus (IS) bezeichnet wird, ruft die zweite SP-Sequenz und die IA hervor.
- Die SPs von S1 und S2 werden automatisch durch die Software bestimmt, die die Marker auf dem ersten und letzten Minimalwert auf den Spuren positioniert, um SP1 und SP2 zu generieren, und gleichzeitig ihre Dauer berechnet. Die IA-Dauer wird zwischen dem letzten Minimalwert von SP1 und dem ersten Minimalwert von SP2 berechnet.
Bei dem getesteten Probanden war der S2-Reiz in der Lage, beide SPs hervorzurufen, während bei einem normalen Probanden der S2-Reiz normalerweise nur den SP1 oder höchstens einen SP2 von reduzierter Dauer hervorrufen kann. Wie in Tabelle 1 gezeigt, wurde festgestellt, dass die Dauer des durch S2 hervorgerufenen SP1 sehr stabil war, ohne signifikante Unterschiede in der Dauer des durch S1 erzeugten SP1 (Δ= -1ms für Ch1 und Δ= -2 ms für Ch2), während die von S2 am rechten und linken Masseter ausgelösten SP1 (61 ms bzw. 54 ms) länger waren als die von S1 ausgelösten (39 ms bzw. 35 ms). Die Unterschiede betrugen +22 ms für Ch1 (rechter Masseter) und +19 ms für Ch2 (linker Masseter). Folglich zeigte die Dauer der IA deutliche Unterschiede zwischen S2 und S1. Die Dauer der durch S2 hervorgerufenen IA betrug 12 ms gegenüber 23 ms des S1-Reizes für den rechten Masseter (Ch1) und 17 ms gegenüber 30 ms des S1-Reizes für den linken Masseter (Ch2) mit einem Unterschied zwischen den Reaktionen, die durch S2 minus S1 hervorgerufen wurden, von -11 ms bzw. -13 ms.
| Tabelle 1 | ||||||
|---|---|---|---|---|---|---|
| Beschreibung der Positionierung und Messungen der Marker
für den Erholungszyklus des Masseter-Inhibitionsreflexes (rcMIR): |
||||||
| S1 | S2 | |||||
| EMG-Spuren | Marker | Anstiegs-Latenz
S1 (ms) |
Marker | Anstiegs-Latenz
S2 (ms) |
||
| 1A | 11 | 13 | 1E | 12 | 12 | |
| Ch1
Right masseter muscle |
1B | 24 | 1F | 24 | ||
| 1C | 47 | 39 | 1G | 37 | 61 | |
| 1D | 86 | 1H | 98 | |||
| 2A | 10 | 16 | 2E | 13 | 14 | |
| Ch2
Left masseter muscle |
2B | 26 | 2F | 27 | ||
| 2C | 56 | 35 | 2G | 44 | 54 | |
| 2D | 91 | 2H | 98 | |||


3 Schritt: Gehirn-MRT (Magnetresonanztomographie)
Die MRT des Gehirns wurde unter Verwendung von Turbo Spin Echo, Fluid Attenuated Inversion Recovery und Gradient Echo Sequenzen durchgeführt, bevor und nach intravenöser Verabreichung eines Kontrastmittels. Die Ergebnisse zeigten das Vorhandensein eines rundlichen Bereichs von etwa 1,5 cm Durchmesser in der Nähe der Vierhügelgrube auf Höhe der Zirbeldrüse. Es gab auch eine leichte Dilatation des supratentoriellen Ventrikelsystems, das in der Achse erschien und besonders in der Nähe der Temporalkrümmung deutlich wurde, mit einem periventrikulären Rand mit einem Transependym-Flüssigkeitsabsorptionsphänomen.[1] Die Signalcharakteristiken der Formation legten eine vorläufige Diagnose eines Pineal-Cavernoms nahe. (Abbildungen 2 und 3)
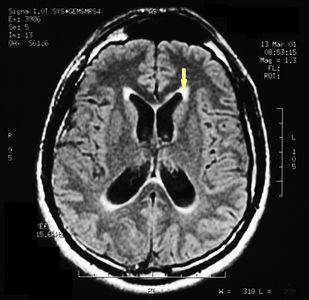
Abbildung 2: Der periventrikuläre Rand, der auf eine Transependymale Flüssigkeitsabsorption hinweist, ist sichtbar (Pfeil).
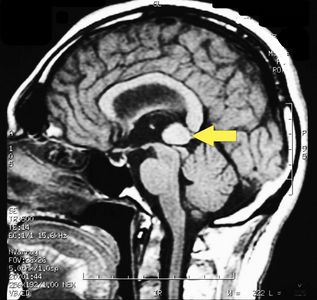
Abbildung 3: Sagittalebene der MRT des Gehirns mit Kontrastmittel (Gadolinium), das umfangreiche Kavernom ist sichtbar (Pfeil).


Abschließende Überlegungen
Wie aus der 'Visuellen kognitiven Galerie' ersichtlich ist, wird der neurologische Kontext durch den Beitrag zur Entschlüsselung der Maschinensprache durch den rcMIR-Test ( ) bereichert, um die Diagnose endgültig mit dem MRT-Bericht ( ) abzuschließen. Das diagnostische Modell Masticationpedia unterstützt nicht nur den Arzt bei komplexen Diagnosen, sondern ist vor allem ein Element zur Umsetzung unseres Grundwissens ( ). Es wird nützlich sein, diese kognitive Interpretation darzustellen, indem sie mit den Bildern der Aussagen des neurologischen Kontexts in Beziehung gesetzt wird.



- Visual Cognitive gallery
Jaw jerk: As a first step, the assertion that the specific phase highlights an anomaly, even if of minor importance, such as amplitude asymmetry, should always be considered, but the absolute value of the amplitudes on each side should also be noted at the same time. Note in the lower section of the window that the right masseter shows an amplitude of 5 mV while the left one of 8.50 mV. The question to ask is: What is the population mean jaw jerk amplitude? This question is of essential importance because it allows us to understand whether this asymmetry is primarily functional or organic and secondly to quantify its absolute response, but with respect to what?

Trigeminale Root-MEPs: Trigeminale Root-MEPs: Die motorisch evozierte Reaktion der trigeminalen Wurzeln, die auf den Massetermuskeln aufgezeichnet wird, wird als bRoot-MEPs bezeichnet..[2][3] Diese neurophysiologische Antwort zeigt die anatomische Komponente des trigeminalen Motoriksystems der untersuchten Person und kontextuell den absoluten Amplitudenwert an, der offensichtlich der neuronalen Energie entspräche, die durch die Depolarisation aller trigeminalen Motorfasern erzeugt wird, die in der motorischen Wurzel enthalten sind. Diese sehr wichtige Information bestimmt den absoluten Wert, auf den sich andere Reflexe wie der Kieferreflex und die laterale Symmetrie beziehen. Bereits aus diesen ersten beiden Fragen können wir schließen, dass die durchschnittliche Amplitude ≊ 30% der Root-MEPs weit über dem Grenzwert liegt, nämlich ≊ 79%, was eine Art Hypererregbarkeit der Mittelhirnreaktion darstellt.
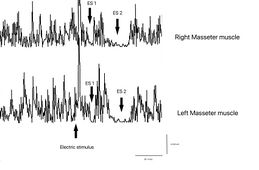
Masseteric Silent Period: Zweifel könnten aufkommen, ob man sich in einer Situation degenerativer und/oder demyelinisierender Neuropathie befindet, aber der Test der elektrischen masseterischen Silent-Periode hebt diese Hypothese auf, da sowohl die Latenz als auch die Dauer der ersten und zweiten Silent-Periode (ES1 und ES2) symmetrisch sind. Auch die dazwischenliegende Aktivität zur Reaktivierung der motorischen Einheiten, die die beiden Silent-Perioden trennt, führt zu Symmetrie im Integralbereich zwischen den Seiten. Diese Daten können nur die Hypererregbarkeit des trigeminalen Systems hervorheben, indem ein zweiter elektrischer Reiz nach dem ersten gekoppelt wird und das beschriebene Phänomen hervorruft, d.h. den Erholungszyklus des Inhibitorischen Masseter-Reflexes (rcRIM).

Recovery Cycle of MIR: Der Erholungszyklus des masseterischen inhibitorischen Reflexes (crRIM) muss konzeptuell mit der übertriebenen Amplitude des Kieferreflexes gekoppelt werden, da dies auf eine Hypererregbarkeit hinweist, die sicherlich die mesenzephale Kerne mit erhöhter Erregbarkeit der γ-Motoneuronen und der polysynaptischen Schaltung umfasst, die für die Hemmung des Motoneurons der masseterischen Silent-Periode verantwortlich ist. Berücksichtigen Sie den Wert des kognitiven Prozesses des neuronalen Netzwerks, das den Entschlüsselungsschlüssel der neuralen Hypererregbarkeit im crRIM identifiziert hat. Nur mit der Kieferreflexuntersuchung hätten wir die Entschlüsselung nie erreicht, bis sich die Symptome des Patienten verschlechtert hätten und eine MRT erforderlich geworden wäre. Wir alle wissen jedoch, dass diese Eventualität verzögert und fatal sein könnte. Logischerweise hätte die Diagnose nach diesem Prozess bereits 10 Jahre früher gestellt werden können, da eine Art abgeschwächter Hypererregbarkeit sicherlich bereits vorhanden gewesen wäre und den Arzt misstrauisch gemacht hätte.

MR imaging of Pineal Cavernoma: MRT-Bildgebung eines Pineal-Cavernoms: Die Schlussfolgerung des bereits beschriebenen Prozesses deutet darauf hin, dass ein Pineal-Cavernom aufgrund seiner anatomischen Position sehr schwer zu behandeln ist. Der Patient wurde von Prof. Albino Bricolo an ein spezialisiertes Zentrum für Neurochirurgie in Verona überwiesen, wo es ihm gelang, die vaskuläre Malformation durch endoskopische Chirurgie zu beseitigen und unserem lieben Patienten 'Bruxer' ein würdevolles Leben ohne eine Zahnschiene zu ermöglichen.









|
| |||||||


Diskussion
Das Hauptziel dieser Studie war die elektrophysiologische Dokumentation der Übererregbarkeit des Trigeminusnervensystems bei einem Patienten mit Zirbeldrüsenkavernom mit ausgeprägten Symptomen von OP und Bruxismus, der gegen jegliche pharmakologische oder odontologische Behandlung resistent war.
Wir fanden Hinweise auf eine Aktivierung und periphere Sensibilisierung der nozizeptiven Fasern, der primären und sekundären nozizeptiven Neuronen im ZNS und der endogenen Schmerzkontrollsysteme, einschließlich sowohl der hemmenden als auch der erleichternden Prozesse bei unserem Probanden.
Die Konzentration von extrazellulärem Glutamat bei 13 Patienten mit kavernösen Angiomen[4] wurde im Vergleich zu physiologischen Konzentrationen als erhöht berichtet. Hohe Glutamatwerte können negative Auswirkungen auf das Gehirn durch exzitotoxische Mechanismen verursachen, einschließlich der Degeneration der oberflächlichen Netzhautschicht bei einer Maus nach wiederholter Verabreichung von Glutamat, was als "Glutamat-Exzitotoxizität"[5] bezeichnet wird und durch eine Hyperaktivierung[6] der NMDA-Rezeptoren entsteht. In einer Studie, in der die Neuronen des Trigeminusganglions KCl ausgesetzt wurden, war die berechnete Freisetzung von Glutamat um das Zehnfache höher als das Basalniveau.[7] Darüber hinaus wurde eine signifikante Reduktion der durch Kalium induzierten Glutamatfreisetzung mit Zugabe von ω-Agatoxin TK, einem starken P/Q-Calciumkanalblocker, beobachtet, während der N-Typ-Calciumkanalblocker ω-Cgtx Conotoxin einen ähnlichen Effekt hatte.[8] Nimodipin, ein L-Typ-Calciumkanalblocker, reduzierte ebenfalls die Menge der durch Kalium induzierten Glutamatfreisetzung.[9] Diese Studien legen nahe, dass die P/Q-, N- und L-Typ-Calciumkanäle jeweils einen signifikanten Anteil der depolarisationsassoziierten Glutamatfreisetzung vermitteln.
Die Freisetzung von Glutamat ist offensichtlich ein viel breiteres und komplexeres Phänomen. NMDA-, Kainat- und AMPA-Ionotroprezeptoren sowie die metabotropen Glutamatrezeptoren wurden in der oberflächlichen Lamina des Nucleus caudalis des Trigeminus bei Mäusen gefunden.[10] NMDA- und AMPA-Rezeptorantagonisten können die Übertragung der nozizeptiven trigemino-vaskulären Signale blockieren [11][12] und den hohen Gehalt an c-fos im Nucleus caudalis des Trigeminus nach zisternaler Injektion von Capsaicin reduzieren.[13] Darüber hinaus führen Mikroinjektionen von ω-Agatoxin in den ventrolateralen Bereich des periaquäduktalen Graus zu einer erleichterten Reaktion der nozizeptiven Aktivität im Trigeminus-Nucleus caudalis (TNC), die durch tonische elektrische Stimulation der supratentorialen parietalen Dura, benachbart zur mittleren Hirnhautarterie, aktiviert wird.[14] Diese Reaktion kann durch antinozizeptive und/oder pronozizeptive Effekte erfolgen, da das Vorhandensein von P/Q-Typ-Calciumkanälen auf synaptischer Ebene für die präsynaptischen Aktionspotentiale erforderlich ist, um mit den Prozessen der Neurotransmitterfreisetzung zu koppeln.[15] Zu beachten ist, dass die präsynaptischen Afferenzen im PAG auf GABAerge hemmende Interneurone und auf absteigende Projektionsneurone positioniert sind. Daher kann der erleichternde Effekt durch eine erhöhte Freisetzung von GABA erklärt werden, die indirekt die Neurone des Hinterhorns enthemmen würde, oder durch einen direkten pronozizeptiven Mechanismus.[16] Diese experimentellen Ergebnisse tragen dazu bei, ein besseres Verständnis der klinischen Manifestationen von Schmerzen und der Hypererregbarkeit des zentralen Nervensystems zu gewinnen, die bei Fällen von zerebralen kavernösen Malformationen gefunden werden.
In der Tat zeigte eine Blinkreflexstudie an einem 38-jährigen Patienten mit rechtsseitigen hemikranialen Symptomen im Zusammenhang mit einem pontinen Kavernom, das den Bereich des Nucleus raphes magnus betrifft, eine Verringerung der Schmerzschwelle und eine anhaltende Erleichterung der R2-Antwort, mit einem Unterschied in der Anfangslatenz von 4,4 ms weniger auf der Seite, die die Symptome zeigte. Dies bestätigt eine regulierende Rolle für die Freisetzung von Neurotransmittern durch den Nucleus raphes magnus, der eine absteigende inhibitorische Kontrolle über den Trigeminus-Nucleus caudalis (TNC)[17] und über den gesamten antinozizeptiven mesenzephalen Komplex aufweist.[18] Unsere Ergebnisse legen eine Hypererregbarkeit des trigeminalen Nervensystems bei unserem Probanden nahe, wie folgt. Zunächst riefen wir eine direkte Reaktion des trigeminalen Motorsystems (bR-MEPs) hervor, um einen Referenzwert und eine Amplitudensymmetrie zu erhalten, da die direkte Reaktion des trigeminalen motorischen Astes nicht von irgendeinem Konditionierungseffekt beeinflusst wurde. Ein Vergleich zwischen den Kieferreflexantworten und den ipsilateralen Reaktionen der R-MEPs zeigte ein viel höheres Amplitudenverhältnis als bei normalen Probanden[19] (Tabelle 1). Diese Daten deuten daher auf das Vorhandensein einer Hypererregbarkeit des trigeminalen Systems hin.
Der erleichternde Effekt auf den Masseterreflex könnte indirekt sein. Die höchste Konzentration von Premotoneuronen in den orofazialen motorischen Kernen findet sich in den bulberen und pontinen retikulären Formationen, die benachbart zu den motorischen Kernen selbst liegen, wo diese GABAerg, glycinerg und glutamatergisch-artige Premotoneuronen sind.[20] Darüber hinaus bestätigt der signifikante Anstieg des SP2-Erholungszyklus von S2 im Vergleich zur Reaktion von S1 (Tabelle 1) die Hypothese einer Hypererregbarkeit des trigeminalen Systems. In einer in vitro durchgeführten Studie an enzephalen Schnitten zeigte die[21] intrazelluläre Aufzeichnung von Interneuronen des peritrigeminalen Bereichs (PeriV), der den trigeminalen motorischen Kern (NVmt) umgibt, und der parvocellulären retikulären Formation (PCRt), dass die elektrische Stimulation der benachbarten Bereiche sowohl exzitatorische postsynaptische Potentiale (EPSPs) als auch inhibitorische postsynaptische Potentiale (IPSPs) hervorrufen konnte. Alle EPSPs, die durch Stimulation des PeriV, PCRt und NVmt induziert wurden, erwiesen sich als empfindlich gegenüber ionotropen Glutamatrezeptorantagonisten (DNQX und APV), während die IPSPs empfindlich auf die GABA- und Glycinrezeptorantagonisten Bicucullin und Strychnin waren. Die Zellen dieser Stichprobe zeigten eine lange Nachhyperpolarisation (AHP).
In einer elektrophysiologischen Studie, die eine Population von Neuronen und Interneuronen im NVmt analysierte,[22] wurden drei Arten von AHP beobachtet: schnell, langsam und biphasisch. Die Mehrheit der Motoneuronen hatte eine schnelle AHP (fAHP), während die meisten Interneuronen eine langsame AHP aufwiesen. Die grundlegenden Eigenschaften dieser Interneuronen ähneln den zuvor beschriebenen "letztordnenden Prämotoneuronen" im PeriV,[23] was darauf hindeutet, dass die Interneuronen im NVmt Teil einer Interneuronenmatrix sind, die den NVmt umgibt, in die die Motoneuronen eingebettet sind. In dieser letzten Studie beschreiben die Autoren die Möglichkeit, wenn auch selten, dass auch Interneurone eine fAHP aufweisen können.
In unserer Studie dringt die erhöhte Dauer des SP2 von S2 in das IA ein, anstatt sich in der EMG-Reaktivierung nach der stillen Periode auszudehnen. Die Afferenzen für den SP2 verlaufen entlang ihres intraaxialen Prozesses entlang des trigeminalen Rückenmarks und verbinden sich mit einer polysynaptischen Kette von erregenden Interneuronen, die sich in der retikulären Formation auf Höhe der pontozerebellären Verbindung befinden. Das letzte Interneuron in der Kette ist hemmend und sendet ipsilaterale und kontralaterale Kollateralen aus, die medial aufsteigen und das rechte und linke spinale trigeminale Komplex erreichen, um die trigeminalen Motoneuronen zu erreichen.[24] Die interneuronale Sensibilisierung im rcMIR könnte mit einer Kombination aus dem exzitatorischen Effekt von Glutamat, einem Beitrag von intraneuronalen fAHP und der Enthemmung der hemmenden Prozesse aufgrund der Wirkung von Glycin und GABA verbunden sein.
Insgesamt legen unsere Daten nahe, dass bestimmte Arten von OP, zumindest solche mit zentraler Ursache, und Bruxismus durch eine Störung und ein Ungleichgewicht der zerebralen Neurobiochemie verursacht werden, insbesondere der exzitatorischen und inhibitorischen Neurotransmitter im trigeminalen Nervensystem.
Dies wirft folgende Fragen auf: Gibt es eine Korrelation zwischen OP und Bruxismus, und kann Bruxismus als klinische Form einer orofazialen Dystonie betrachtet werden?
Hinsichtlich der Korrelation sollte eine Unterscheidung zwischen zentralen und peripheren OP auf der Grundlage der Anamnese und der klinischen Untersuchung erfolgen. Das Muskeldyskomfort des Bruxismus ist hauptsächlich ein peripheres Phänomen, das aus einer Muskelhyperfunktion resultiert, die zur Zerstörung der Myofibrillen und Freisetzung von algogenen Substanzen, einschließlich Myoglobin, in den Blutkreislauf führt. Im Gegensatz dazu könnte bei OP, die sich auf ein oder mehrere Bereiche des Gesichts erstrecken und mit einer klaren Manifestation von nächtlichem oder täglichem Bruxismus korreliert sind, eine zentralisiertere Störung betrachtet werden. In diesen Fällen sind trigeminale elektrophysiologische Untersuchungen besonders informativ, insbesondere der rcMIR, der Blinkreflex, der JJr und der bR-MEPs, um eine Differentialdiagnose zwischen organischen Läsionen des ZNS und funktionellen Erkrankungen wie TMDs durchzuführen.
Daher können Bruxismus und zentrale OP zwar gemeinsam auftreten, sind aber zwei unabhängige Symptome, weshalb viele experimentelle und klinische Studien zu uneindeutigen Schlussfolgerungen gelangen.[25]
Es ist auch möglich, dass Bruxismus eine klinische Form von Dystonie sein kann. Unsere Daten deuten darauf hin, dass Bruxismus eine klinische Manifestation sein kann, die mit einem Ungleichgewicht der Neurotransmitter im ZNS verbunden ist, und daher als subklinischer Zustand einer orofazialen Dystonie oder eines dystonischen Syndroms betrachtet werden sollte. Dennoch tritt dieses Phänomen auch in transitorischer Form bei Kindern auf und löst sich mit dem Durchbruch der Mischzahnung..[26][27]
Viele Studien und diagnostische Forschungsprotokolle, einschließlich der Forschungskriterien für diagnostische Kriterien (RDC), tauchen weiterhin auf dem Gebiet der OP und TMDs auf, obwohl noch kein klarer Konsens innerhalb der internationalen wissenschaftlichen Gemeinschaft erreicht wurde.[28] Die RDC sollte den Patienten als von einem schmerzhaften Syndrom betroffen betrachten und sollte dazu neigen, eine Differentialdiagnose zwischen organischen und/oder funktionellen Pathologien zu stellen.[29]
- ↑ Peter H Yang, Alison Almgren-Bell, Hongjie Gu, Anna V Dowling, Sangami Pugazenthi, Kimberly Mackey, Esther B Dupépé, Jennifer M Strahle. Etiology- and region-specific characteristics of transependymal cerebrospinal fluid flow. J Neurosurg Pediatr. 2022 Aug 12;1-11. doi: 10.3171/2022.7.PEDS2246. Online ahead of print.
- ↑ Frisardi G. The use of transcranial stimulation in the fabrication of an occlusal splint. J Prosthet Dent, 1992, DOI: 10.1016/0022-3913(92)90345-b
- ↑ G. Frisardi 1, P. Ravazzani, G. Tognola, F Grandori. Electric versus magnetic transcranial stimulation of the trigeminal system in healthy subjects. Clinical applications in gnathology. J Oral Rehabil.1997 Dec;24(12):920-8. doi: 10.1046/j.1365-2842.1997.00577.x.
- ↑ von Essen C, Rydenhag B, Nystrom B, Mozzi R, van Gelder N, Hamberger A. High levels of glycine and serine as a cause of the seizure symptoms of cavernous angiomas? J Neurochem. 1996;67(1):260–264. [PubMed] [Google Scholar]
- ↑ Lau A, Tymianski M. Glutamate receptors, neurotoxicity and neurodegeneration. Pflugers Arch. 2010;460(2):525–542. doi: 10.1007/s00424-010-0809-1. [PubMed] [CrossRef] [Google Scholar]
- ↑ Meldrum B, Garthwaite J. Excitatory amino acid neurotoxicity and neurodegenerative disease. Trends Pharmacol Sci. 1990;11(9):379–387. doi: 10.1016/0165-6147(90)90184-A. [PubMed] [CrossRef] [Google Scholar]
- ↑ Xiao Y, Richter JA, Hurley JH. Release of glutamate and CGRP from trigeminal ganglion neurons: role of calcium channels and 5-HT1 receptor signaling. Mol Pain. 2008;4:12. doi: 10.1186/1744-8069-4-12. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
- ↑ McCleskey EW, Fox AP, Feldman DH, Cruz LJ, Olivera BM, Tsien RW, Yoshikami D. Omega-conotoxin: direct and persistent blockade of specific types of calcium channels in neurons but not muscle. Proc Natl Acad Sci U S A. 1987;84(12):4327–4331. doi: 10.1073/pnas.84.12.4327. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
- ↑ Hockerman GH, Johnson BD, Abbott MR, Scheuer T, Catterall WA. Molecular determinants of high affinity phenylalkylamine block of L-type calcium channels in transmembrane segment IIIS6 and the pore region of the alpha1 subunit. J Biol Chem. 1997;272(30):18759–18765. doi: 10.1074/jbc.272.30.18759. [PubMed] [CrossRef] [Google Scholar]
- ↑ Tallaksen-Greene SJ, Young AB, Penney JB, Beitz AJ. Excitatory amino acid binding sites in the trigeminal principal sensory and spinal trigeminal nuclei of the rat. Neurosci Let. 1992;141(1):79–83. doi: 10.1016/0304-3940(92)90339-9. [PubMed] [CrossRef] [Google Scholar]
- ↑ Storer RJ, Goadsby PJ. Trigeminovascular nociceptive transmission involves N-methyl-D-aspartate and non-N-methyl-D-aspartate glutamate receptors. Neuroscience. 1999;90(4):1371–1376. doi: 10.1016/S0306-4522(98)00536-3. [PubMed] [CrossRef] [Google Scholar]
- ↑ Goadsby PJ, Classey JD. Glutamatergic transmission in the trigeminal nucleus assessed with local blood flow. Brain Res. 2000;875(1–2):119–124. [PubMed] [Google Scholar]
- ↑ Waeber C, Moskowitz MA, Cutrer FM, Sanchez Del Rio M, Mitsikostas DD. The NMDA receptor antagonist MK-801 reduces capsaicin-induced c-fos expression within rat trigeminal nucleus caudalis. Pain. 1998;76(1–2):239–248. [PubMed] [Google Scholar]
- ↑ Knight YE, Bartsch T, Kaube H, Goadsby PJ. P/Q-type calcium-channel blockade in the periaqueductal gray facilitates trigeminal nociception: a functional genetic link for migraine? J Neurosci. 2002;22(5):RC213. [PMC free article] [PubMed] [Google Scholar]
- ↑ Dunlap K, Luebke JI, Turner TJ. Exocytotic Ca2+ channels in mammalian central neurons. Trends Neurosci. 1995;18(2):89–98. doi: 10.1016/0166-2236(95)93882-X. [PubMed] [CrossRef] [Google Scholar]
- ↑ Pan ZZ, Williams JT, Osborne PB. Opioid actions on single nucleus raphe magnus neurons from rat and guinea-pig in vitro. J Physiol. 1990;427:519–532. [PMC free article] [PubMed] [Google Scholar]
- ↑ Hentall ID. Interactions between brainstem and trigeminal neurons detected by cross-spectral analysis. Neuroscience. 2000;96(3):601–610. doi: 10.1016/S0306-4522(99)00593-X. [PubMed] [CrossRef] [Google Scholar]
- ↑ Jiang M, Behbehani MM. Physiological characteristics of the projection pathway from the medial preoptic to the nucleus raphe magnus of the rat and its modulation by the periaqueductal gray. Pain. 2001;94(2):139–147. doi: 10.1016/S0304-3959(01)00348-7. [PubMed] [CrossRef] [Google Scholar]
- ↑ Cruccu G, Berardelli A, Inghilleri M, Manfredi M. Functional organization of the trigeminal motor system in man. A neurophysiological study. Brain. 1989;112(5):1333–1350. doi: 10.1093/brain/112.5.1333. [PubMed] [CrossRef] [Google Scholar]
- ↑ Li YQ, Takada M, Kaneko T, Mizuno N. GABAergic and glycinergic neurons projecting to the trigeminal motor nucleus: a double labeling study in the rat. J Comp Neurol. 1996;373(4):498–510. doi: 10.1002/(SICI)1096-9861(19960930)373:4<498::AID-CNE3>3.0.CO;2-X. [PubMed] [CrossRef] [Google Scholar]
- ↑ Bourque MJ, Kolta A. Properties and interconnections of trigeminal interneurons of the lateral pontine reticular formation in the rat. J Neurophys. 2001;86(5):2583–2596. [PubMed] [Google Scholar]
- ↑ McDavid S, Verdier D, Lund JP, Kolta A. Electrical properties of interneurons found within the trigeminal motor nucleus. Eur J Neurosci. 2008;28(6):1136–1145. doi: 10.1111/j.1460-9568.2008.06413.x. [PubMed] [CrossRef] [Google Scholar]
- ↑ Kolta A, Westberg KG, Lund JP. Identification of brainstem interneurons projecting to the trigeminal motor nucleus and adjacent structures in the rabbit. J Chem Neuroanat. 2000;19(3):175–195. doi: 10.1016/S0891-0618(00)00061-2. [PubMed] [CrossRef] [Google Scholar]
- ↑ Ongerboer de Visser BW, Cruccu G, Manfredi M, Koelman JH. Effects of brainstem lesions on the masseter inhibitory reflex. Functional mechanisms of reflex pathways. Brain. 1990;113(3):781–792. doi: 10.1093/brain/113.3.781. [PubMed] [CrossRef] [Google Scholar]
- ↑ Svensson P, Jadidi F, Arima T, Baad-Hansen L, Sessle BJ. Relationships between craniofacial pain and bruxism. J Oral Rehabil. 2008;35(7):524–547. doi: 10.1111/j.1365-2842.2008.01852.x. [PubMed] [CrossRef] [Google Scholar]
- ↑ Watts MW, Tan EK, Jankovic J. Bruxism and cranial-cervical dystonia: is there a relationship? Cranio. 1999;17(3):196–201. [PubMed] [Google Scholar]
- ↑ Monaco A, Ciammella NM, Marci MC, Pirro R, Giannoni M. The anxiety in bruxer child. A case–control study. Minerva Stomatol. 2002;51(6):247–250. [PubMed] [Google Scholar]
- ↑ Lobbezoo F, Visscher CM, Naeije M. Some remarks on the RDC/TMD Validation Project: report of an IADR/Toronto-2008 workshop discussion. J Oral Rehabil. 2010;37(10):779–783. doi: 10.1111/j.1365-2842.2010.02091.x. [PubMed] [CrossRef] [Google Scholar]
- ↑ Frisardi G, Chessa G, Sau G, Frisardi F. Trigeminal electrophysiology: a 2 × 2 matrix model for differential diagnosis between temporomandibular disorders and orofacial pain. BMC Musculoskelet Disord. 2010;11:141. doi: 10.1186/1471-2474-11-141. [PMC free article] [PubMed] [CrossRef] [Google Scholar]