Difference between revisions of "The mysterious "muscle tone""
| (One intermediate revision by the same user not shown) | |||
| Line 1: | Line 1: | ||
Se leggiamo attentamente sia le motivazioni del Research Diagnostic Criteria propenso ad eliminare le procedure cliniche di 'Stimolazione Elettrica Transcutanea' a fronte delle considerazioni di efficacia terapeutica e di supporto clinico nelle riabilitazioni del sistema masticatorio dei lavori sperimentali presentati nei capitoli precedenti non si può che rimanere 'basiti' perchè la Scienza, con tutte le sue dinamicità e rivoluzioni interne non può generare, comunque, confusione epistemologica causando a valle difficoltà ed errori diagnostici. | |||
Per questo motivo pensiamo sia dovuto un po di chiarezza. Il primo sospettato è la 'Interdisciplinarità' intesa come la conoscenza della scienza al di fuori della propria specializzazione, infatto, in neurofisiologia il tomo muscolare non si rileva con un esame EMG di superficie. Riportiamo un esempio clinico per essere più chiari nell'asserzione. | |||
In figura 1 viene riportato una serie di tracciati EMG di una paziente sofferente di algie orofacciali con particolare localizzazione nel massetere di sinistra ( per maggiori dettagli vedi capitoloXXXX). Il quadro elettromiografico dei masseteri è stato determinato seguendo una logica sequenza riportata nella figura 1. Come si può osservare, l’asimmetria di lato dell’attività EMG con elettrodi di superficie dei masseteri in stato di rilassamento (fig.1A) con la mandibola mantenuta in posizione di riposo era tale da richiedere una EMG ad ago del massetere sinistro. L’attività registrata con questa tecnica (fig.1B) ha mostrato una scarica con frequenza stabile di 20 Hz il che presuppose uno studio dell’unità motoria. Lo studio delle unità motorie del massetere sinistro (fig.1C), attraverso ago coassiale, ha automaticamente selezionato 26 unità motorie di cui sono state analizzate la forma, la durata, gli spikes ed i turns di ciascuna unità. I dati sono riportati nella tabella (fig.1D) Statisticamente si possono rilevare i seguenti parametri: ampiezza media di <math>\approxeq348\mu V</math>, una durata di 8.7 mS., il 23% di unità polifasiche. Questo quadro clinico rappresenta il tipico fenomeno fisiopatologico in cui il paziente riferisce dolore ma molto spesso la diagnosi rimane “difficoltà nel rilassamento muscolare”, “ dolore orofacciale atipico” o ancor meglio “fibromialgia” e di conseguenza la terapia farmacologica rimane quella sintomatica. <gallery widths="350" heights="200" perrow="2" slideshow""=""> | |||
File:Distonic 2.jpg|'''Figura 1a:''' EMG di superficie che mostra una ampia attività di scarica delle unità motorie (MUs) nel muscolo massetere di sinistra | |||
File:Distonic 3.jpg|'''Figura 1b:''' Analisi della frequenza di scarica delle unità motorie masseteri (MUs) | |||
File:Distonic 1.jpg|'''Figura 1c:''' Analisi della morfologia della Unità motoria masseteri (MUs) | |||
File:Distonic 4.jpg|'''Figura 1d:''' Calcolo dei parametri di scarica delle MUs | |||
</gallery> | |||
Quello che inconfutabilmente evoca questo paragrafo da un punto di vista scientifico è che nel campo odontoiatrico, troppo concentrato nelle proprie dogmi specialistici, si interpreta l'attività EMG a riposocome tono muscolare e da qui si giunge alla eventuale diagnosi di ipertono e contestualmente trattamento con TENS. Se osserviamo attentamente la figura Ia | |||
[[File:Distonic 2.jpg|border|center|500x500px]] | |||
Lo studio di Edward Stanek IV<ref>Edward Stanek 4th 1, Erica Rodriguez 1, Shengli Zhao 1, Bao-Xia Han 1, Fan Wang 2. Supratrigeminal Bilaterally Projecting Neurons Maintain Basal Tone and Enable Bilateral Phasic Activation of Jaw-Closing Muscles. J Neurosci. 2016 Jul 20;36(29):7663-75. | |||
doi: 10.1523/JNEUROSCI.0839-16.2016.</ref> esplora il ruolo dei neuroni bilaterali proiettanti (BPN) del nucleo sopratrigeminale (SupV) nel controllo del tono basale e dell'attivazione fasica dei muscoli mandibolari, concentrandosi sui muscoli coinvolti nella chiusura della mandibola. Il documento sottolinea come i BPN possano mediare sia un'attivazione bilaterale fasica dei muscoli masseteri durante i movimenti di chiusura mandibolare, sia mantenere un livello di tono muscolare bilanciato e ridotto attraverso un'inibizione tonica. | |||
== Introduzione == I muscoli mandibolari sono fondamentali per numerosi comportamenti orofacciali, inclusi la masticazione, la respirazione e il parlare. Tuttavia, le disfunzioni in queste attività possono portare a condizioni patologiche come il bruxismo e i disturbi dell'articolazione temporo-mandibolare<ref>Balasubramaniam R, Ram S. 2008</ref>. Un elemento cruciale per il controllo della mandibola è la coordinazione bilaterale, che richiede un'attivazione simultanea e sincronizzata dei muscoli mandibolari su entrambi i lati. Il SupV e i nuclei reticolari parvicellulari (Rt-MoVII), situati dorsalmente rispetto al nucleo motore facciale, contengono BPN che proiettano simultaneamente ai motoneuroni mandibolari bilaterali<ref>Li et al., 1993</ref>. Il SupV, in particolare, svolge un ruolo chiave nel mediare riflessi orofacciali e sensoriali, come dimostrato da studi precedenti, che ne hanno evidenziato il coinvolgimento nell'inibizione motoria dei riflessi mandibolari dolorifici<ref>Kamogawa et al., 1994</ref><ref>Minkels et al., 1995</ref>. | |||
Questo studio mira a caratterizzare i BPN di SupV e Rt-MoVII, identificandone la neurochimica, i percorsi di proiezione e il ruolo funzionale nei movimenti mandibolari, utilizzando tecniche avanzate di manipolazione genetica e optogenetica<ref>Stanek et al., 2014</ref>. | |||
== Metodi == Per raggiungere questo obiettivo, il team di ricerca ha combinato tecniche di tracciamento virale retrogrado con il sistema split-Cre, consentendo di isolare selettivamente e manipolare i BPN<ref>Wang et al., 2012</ref>. L'espressione di proteine optogenetiche come Channelrhodopsin-2 (ChR2) ha permesso l'attivazione controllata dei BPN, mentre l'uso della tossina tetanica (TeNT) ha consentito di silenziarli in modo permanente, rivelando le conseguenze funzionali dell'inattività di questi neuroni sui movimenti della mandibola. | |||
L'ibridazione in situ ha dimostrato che la popolazione di BPN è mista, contenendo neuroni eccitatori (glutammatergici) e inibitori (GABAergici e glicinergici)<ref>Travers et al., 2005</ref><ref>Paik et al., 2009</ref>. Le proiezioni di questi neuroni non si limitano ai nuclei motori trigeminali, ma coinvolgono anche altri gruppi motori orofacciali e regioni mesencefaliche implicate nelle funzioni motorie<ref>Fay e Norgren, 1997</ref><ref>Kolta et al., 2000</ref>. | |||
== Risultati == === Funzioni di SupV e Rt-MoVII === I BPN di SupV sono fondamentali per il mantenimento del tono basale del muscolo massetere, come evidenziato dal fatto che il loro silenziamento provoca un aumento significativo del tono muscolare bilaterale, suggerendo che questi neuroni esercitano un'inibizione tonica sui motoneuroni mandibolari. Questo effetto è mediato da input inibitori GABAergici e glicinergici, che regolano il livello di attività dei motoneuroni<ref>Brooks e Peever, 2008</ref><ref>Gemba-Nishimura et al., 2010</ref>. Tuttavia, i BPN eccitatori di SupV giocano anche un ruolo cruciale durante i movimenti fasici della mandibola, come quelli coinvolti nella masticazione, in cui l'attivazione dei BPN induce scoppi sincronizzati di attivazione muscolare bilaterale<ref>Inoue et al., 1994</ref>. | |||
Rt-MoVII, al contrario, sembra avere un ruolo minore nella regolazione diretta dei movimenti di chiusura mandibolare. Nonostante la presenza di BPN in quest'area, il silenziamento di Rt-MoVII non ha prodotto effetti significativi sull'attivazione muscolare, suggerendo che i circuiti paralleli possono compensare per la loro funzione<ref>Yamamoto et al., 1989</ref><ref>Inoue et al., 1992</ref>. | |||
=== Manipolazione Optogenetica e Silenziamento Cronico === Gli esperimenti di attivazione optogenetica mediante ChR2 hanno rivelato che l'attivazione dei BPN eccitatori di SupV può sincronizzarsi con i movimenti naturali di chiusura della mandibola, amplificando l'attività muscolare<ref>Madisen et al., 2012</ref>. In particolare, l'attivazione di questi neuroni durante periodi di riposo ha provocato scoppi di attivazione muscolare simili a quelli osservati durante la masticazione. | |||
D'altra parte, il silenziamento cronico dei BPN con TeNT non ha indotto cambiamenti immediati nel tono muscolare basale, suggerendo che i meccanismi compensatori possono attenuare gli effetti dell'inibizione cronica, riducendo l'impatto del silenziamento dei BPN a lungo termine<ref>Yamamoto et al., 2003</ref>. | |||
== Discussione == I risultati dello studio supportano l'idea che i BPN di SupV esercitino un duplice controllo sull'attivazione muscolare mandibolare: da un lato, essi sono responsabili di mantenere un tono muscolare bilanciato attraverso un'inibizione tonica; dall'altro, consentono attivazioni fasiche bilaterali, come quelle necessarie per la masticazione<ref>Travers et al., 2005</ref>. Il bilanciamento tra questi due input, eccitatorio e inibitorio, è cruciale per garantire un funzionamento normale della mandibola e prevenire disfunzioni muscolari che possono portare a condizioni patologiche come il bruxismo e altri disturbi orofacciali<ref>Richter e Löscher, 1998</ref>. | |||
In sintesi, i BPN di SupV sono una popolazione mista di neuroni eccitatori e inibitori, che regolano finemente i movimenti della mandibola e l'attività dei muscoli orofacciali. L'attivazione optogenetica dimostra il loro ruolo nel generare scoppi di attivazione muscolare sincronizzati con i movimenti naturali della mandibola, mentre il silenziamento cronico e optogenetico rivela il loro contributo al mantenimento di un tono muscolare appropriato. Saranno necessari studi futuri per comprendere come queste reti neuronali siano influenzate da input sensoriali e centrali, e come i BPN contribuiscano ai disturbi orofacciali associati al sonno e ai movimenti orali alterati. | |||
---- | ---- | ||
===5° Clinical case: Spontaneous Electromyographic Activity=== | ---- | ||
===5° Clinical case: Spontaneous Electromyographic Activity === | |||
Paziente femmina di età 65 anni che riferisce principalmente Dolore Orofacciale (OP)nell'emirato sinistro della faccia ed in particolare un dolore che si irradia dai masseteri alla TMJ ed al muscolo temporale di sinistra. Dopo circa 2 anni da un episodio di perdita improvvisa di coscienza al momento in cui il proprio dentista eseguì una trocleare per paradontologia dell'emigrata inferiore sinistra. Da quel momento iniziarono dolori improvvisi di tipo untorio e poi diffusi a tutta l'emifaccia sinistra anche nella masticazione. I colleghi visto la correlazione con la masticazione ed analizzata seguendo il protocollo RDC definirono la paziente affetta da Temporomandibular Disorders (TMDs) | Paziente femmina di età 65 anni che riferisce principalmente Dolore Orofacciale (OP)nell'emirato sinistro della faccia ed in particolare un dolore che si irradia dai masseteri alla TMJ ed al muscolo temporale di sinistra. Dopo circa 2 anni da un episodio di perdita improvvisa di coscienza al momento in cui il proprio dentista eseguì una trocleare per paradontologia dell'emigrata inferiore sinistra. Da quel momento iniziarono dolori improvvisi di tipo untorio e poi diffusi a tutta l'emifaccia sinistra anche nella masticazione. I colleghi visto la correlazione con la masticazione ed analizzata seguendo il protocollo RDC definirono la paziente affetta da Temporomandibular Disorders (TMDs) | ||
| Line 116: | Line 57: | ||
==== Steps EMG ad ago==== | ====Steps EMG ad ago ==== | ||
L’esame EMG dei muscoli scheletrici consiste di quattro step: | L’esame EMG dei muscoli scheletrici consiste di quattro step: | ||
# Attività di inserzione quando inserito l’elettrodo ad ago nel muscolo | #Attività di inserzione quando inserito l’elettrodo ad ago nel muscolo | ||
#Attività spontanea quando il muscolo si trova in condizioni di riposo | #Attività spontanea quando il muscolo si trova in condizioni di riposo | ||
#Potenziali di unità motoria evocate da isolate scariche motorie durante una moderata contrazione volontaria | #Potenziali di unità motoria evocate da isolate scariche motorie durante una moderata contrazione volontaria | ||
#Reclutamento o pattern interferenziale durante progressivo livello di contrazione | # Reclutamento o pattern interferenziale durante progressivo livello di contrazione | ||
=====Attività di inserzione===== | =====Attività di inserzione===== | ||
| Line 142: | Line 83: | ||
Il numero di unità polifasiche aumentano sia nelle miopatie, neuropatie o nelle patologie del motoneurone. La polifasia indica, dunque, una dispersione temporale dei potenziali di fibra muscolari dentro una unità motoria. In alcune anormalità chiamate doppiette o triplette una unità motoria spara due o tre volte ad una intervallo di tempo molto corto e sono rappresentative di un disturbo metabolico associato ad ipereccitabilità del pool motoneurale. | Il numero di unità polifasiche aumentano sia nelle miopatie, neuropatie o nelle patologie del motoneurone. La polifasia indica, dunque, una dispersione temporale dei potenziali di fibra muscolari dentro una unità motoria. In alcune anormalità chiamate doppiette o triplette una unità motoria spara due o tre volte ad una intervallo di tempo molto corto e sono rappresentative di un disturbo metabolico associato ad ipereccitabilità del pool motoneurale. | ||
In figura 5 possiamo osservare un tipico tracciato a minima attività volontaria di MUAP polifasico ed una doppietta che rappresenta uno stato di patologia del motoneurone. Confrontando questa registrazione di unità motoria patologica con alcune della fig.1C, ed in particolare la 5,7,13 e 23 con i valori di ampiezza rispettivi <math>678\mu V;419\mu V3;686\mu V e 530\mu V</math> durata <math>8.2, 4.2,6.2 msec </math> e e 8.4 <math>8.4 msec </math> possiamo affermare che l’attività registrata sul massetere di sinistra, del caso clinico in questione, non ha nessuna caratteristica elettrofisiologica sovrapponibile ad un quadro di danno del II motoneurone. | In figura 5 possiamo osservare un tipico tracciato a minima attività volontaria di MUAP polifasico ed una doppietta che rappresenta uno stato di patologia del motoneurone. Confrontando questa registrazione di unità motoria patologica con alcune della fig.1C, ed in particolare la 5,7,13 e 23 con i valori di ampiezza rispettivi <math>678\mu V;419\mu V3;686\mu V e 530\mu V</math> durata <math>8.2, 4.2,6.2 msec </math> e e 8.4 <math>8.4 msec </math> possiamo affermare che l’attività registrata sul massetere di sinistra, del caso clinico in questione, non ha nessuna caratteristica elettrofisiologica sovrapponibile ad un quadro di danno del II motoneurone. | ||
==Sintesi== | |||
=== 5° Clinical case: Spontaneous Electromyographic Activity === Il 5° caso clinico riguarda una paziente femmina di 65 anni che lamenta dolore orofacciale (OP) nell'emirato sinistro della faccia, descritto come un dolore irradiato dai masseteri fino all'articolazione temporomandibolare (TMJ) e al muscolo temporale sinistro. Il dolore si è manifestato circa due anni dopo un episodio di perdita improvvisa di coscienza verificatosi durante una procedura paradontologica presso il suo dentista, che ha eseguito una trocleare sull'emiarcata inferiore sinistra. Da quel momento, la paziente ha iniziato a soffrire di dolori improvvisi e diffusi su tutta l'emifaccia sinistra, che si acuiscono durante la masticazione. | |||
I colleghi, notando la correlazione con la masticazione e seguendo il protocollo Research Diagnostic Criteria (RDC), diagnosticarono alla paziente un disturbo temporomandibolare (TMDs) <ref>Balasubramaniam R, Ram S. 2008</ref>. | |||
Alla nostra attenzione, abbiamo eseguito tutti i test gnatologici (assiografia, immagini delle articolazioni temporomandibolari, e elettromiografia di superficie) che non indicavano chiaramente la presenza di un TMDs, ma piuttosto un quadro neurologico. In particolare, l'elettromiografia (EMG) di superficie ha mostrato risultati anomali. Il tracciato elettromiografico dei masseteri è stato esaminato seguendo una logica precisa riportata in figura 1. | |||
Come si osserva in figura 1a, l'EMG di superficie del massetere sinistro in condizioni di rilassamento con la mandibola in posizione di riposo ha evidenziato un'attività asimmetrica rispetto al lato destro. Questa asimmetria ha reso necessario eseguire un'EMG ad ago sul massetere sinistro per un'analisi più approfondita. L'elettromiografia ad ago (EMG) ha rivelato una scarica con una frequenza stabile di 20 Hz (figura 1b), suggerendo la necessità di uno studio dell'unità motoria. | |||
L'analisi delle unità motorie del massetere sinistro (figura 1c) ha selezionato automaticamente 26 unità motorie, di cui sono state analizzate la forma, la durata, gli spike e i turns. I dati sono riassunti in figura 1d e mostrano un'ampiezza media di circa 348 μV, una durata di 8.7 ms, e il 23% di unità polifasiche. | |||
Questo quadro clinico riflette un fenomeno fisiopatologico tipico in cui la paziente riferisce dolore, ma la diagnosi rimane spesso vaga, classificata come "difficoltà nel rilassamento muscolare," "dolore orofacciale atipico" o "fibromialgia." Di conseguenza, la terapia è generalmente sintomatica. Tuttavia, tali condizioni dovrebbero spingere il medico ad approfondire le indagini, eseguendo un'analisi EMG ad ago prima di indirizzare la paziente a uno specialista neurologo. | |||
<gallery widths="350" heights="200" perrow="2" slideshow=""> File:Distonic 2.jpg|'''Figura 1a:''' EMG di superficie che mostra una ampia attività di scarica delle unità motorie (MUs) nel muscolo massetere di sinistra File:Distonic 3.jpg|'''Figura 1b:''' Analisi della frequenza di scarica delle unità motorie masseteri (MUs) File:Distonic 1.jpg|'''Figura 1c:''' Analisi della morfologia della Unità motoria masseteri (MUs) File:Distonic 4.jpg|'''Figura 1d:''' Calcolo dei parametri di scarica delle MUs </gallery> | |||
==== Steps EMG ad ago ==== L'esame EMG dei muscoli scheletrici consiste in quattro fasi fondamentali: | |||
Attività di inserzione al momento dell'inserimento dell'elettrodo nel muscolo. | |||
Attività spontanea quando il muscolo è in condizioni di riposo. | |||
Potenziali di unità motoria evocati da scariche isolate durante una moderata contrazione volontaria. | |||
Reclutamento o pattern interferenziale durante un progressivo livello di contrazione. | |||
===== Attività di inserzione ===== In un soggetto normale, l'attività di inserzione si presenta come spike ad alta frequenza, sia positivi che negativi, e rappresenta una risposta a stimolazioni meccaniche dovute all'inserimento dell'ago nel muscolo. Nel caso della nostra paziente, questa attività si è manifestata con una durata di 80 ms ed è stata considerata normale. | |||
Da notare che nei muscoli normali a riposo, non c'è alcuna attività elettrica significativa ad eccezione della regione della placca neuromuscolare, che può manifestare spike di bassa ampiezza (<10-50 μV) e di breve durata (1-2 ms) <ref>Axelsson J, Thesleff S. 1957</ref>. Nel nostro caso clinico, l'assenza di attività di placca sul massetere destro può essere attribuita alla registrazione con elettrodi di superficie, che possono attenuare il segnale <ref>Miledi R. 1960</ref>. Tuttavia, l'attività sul massetere sinistro registrata con elettrodi di superficie ha mostrato un'ampiezza di circa 100 μV, suggerendo un'origine diversa dall'attività di placca, come evidenziato in figura 1b. | |||
===== Attività spontanea ===== [[File | |||
a punta+.jpeg|thumb|'''Figura 2:''' Attività spontanea con scariche a punta positive in un muscolo denervato]] L'attività spontanea nei muscoli denervati è comunemente osservata nelle prime settimane dopo la denervazione e può essere spiegata da un aumento della sensibilità all'acetilcolina (ACh). Questo fenomeno, noto come "ipersensibilità da denervazione," può causare scariche spontanee delle fibre muscolari in risposta a stimoli minimi di ACh<ref>Thesleff S. 1982</ref>. | |||
Nel nostro caso, tuttavia, l'attività spontanea osservata nel massetere sinistro della paziente non presentava le caratteristiche tipiche della denervazione, come le onde a punta positiva o la fibrillazione, come illustrato in figura 2. Questi risultati hanno suggerito che l'attività spontanea registrata non fosse correlata a una patologia del motoneurone inferiore. | |||
===== Potenziali di unità motoria ===== Le unità motorie vengono definite in base all'ampiezza, durata, rise time, e numero di fasi. Un operatore esperto può selezionare potenziali di unità motoria con un rise time di circa 500 μsec per garantire la vicinanza alla sorgente. Nei muscoli facciali, l'ampiezza normale varia tra alcuni centinaia di μV fino a pochi mV, con una durata media tra 5 e 10 ms <ref>Buchthal F. 1957</ref>. In condizioni normali, si osserva una certa presenza di potenziali polifasici (potenziali con quattro o più fasi), ma il loro numero tende ad aumentare in caso di neuropatie o miopatie <ref>Partanen JV, Nousiainen U. 1983</ref>. | |||
La figura 5 mostra un tracciato di attività polifasica, in cui è visibile la presenza di potenziali polifasici e doppiette. Confrontando questi risultati con le registrazioni delle unità motorie della paziente (figura 1c), possiamo escludere la presenza di una patologia del motoneurone inferiore nel caso clinico in esame. | |||
===== Fascicolazioni e scariche ripetitive ===== [[File | |||
.jpeg|thumb|'''Figura 4:''' Fascicolazioni del muscolo orbicolaris oculi sinistro.]] Le fascicolazioni sono contrazioni spontanee di unità motorie e sono comunemente osservate in patologie delle cellule delle corna anteriori, radicolopatie, neuropatie da intrappolamento, e nella sindrome delle fascicolazioni dolorose<ref>Sindermann F, Conrad B. 1973</ref>. Le scariche ripetitive della stessa unità motoria avvengono in burst regolari e sono rappresentative di disturbi del motoneurone. Come si vede in figura 4, le fascicolazioni nel muscolo orbicolare dell'occhio sinistro mostrano una morfologia differente rispetto ai tracciati EMG del massetere sinistro della paziente, confermando l'assenza di patologie da denervazione. | |||
In conclusione, il caso clinico ha dimostrato come l'elettromiografia ad ago sia uno strumento essenziale per identificare anomalie neuromuscolari. Sebbene inizialmente il caso fosse stato diagnosticato come un disturbo temporomandibolare, l'analisi elettromiografica ha rivelato un quadro clinico più complesso, suggerendo l'importanza di una diagnosi neurologica approfondita in pazienti con dolore orofacciale atipico. | |||
---- | |||
==Bruxer== | |||
[[File:Bruxer SP2.jpg|left|200x200px]] | [[File:Bruxer SP2.jpg|left|200x200px]] | ||
Pochi studi hanno tentato di caratterizzare il dolore orofacciale (OP) associato al bruxismo (esaminare le caratteristiche neurobiologiche e fisiologiche dei muscoli mandibolari). Alcuni casi clinici e studi su piccola scala suggeriscono che alcuni farmaci legati ai sistemi dopaminergico, serotoninergico e adrenergico possono sopprimere o esacerbare il bruxismo. Inoltre, la maggior parte di questi studi farmacologici indica che varie classi di farmaci possono influenzare l'attività muscolare correlata al bruxismo, senza esercitare alcun effetto sullo OP. Pertanto, la sensibilizzazione del sistema nocicettivo del trigemino e l'effetto facilitante sui riflessi da stiramento mandibolare e sull'ipereccitabilità neurale sono fenomeni neurofisiopatogenetici correlabili al dolore nella regione craniofacciale. Tuttavia, fino ad ora, non è stata segnalata alcuna correlazione tra OP, disfunzione dei nuclei mesencefalici e facilitazione della nocicezione trigeminale, ad eccezione di uno studio clinico su un paziente affetto da cavernoma pineale, che ha evidenziato una relativa facilitazione del sistema nocicettivo trigeminale attraverso i riflessi. | Pochi studi hanno tentato di caratterizzare il dolore orofacciale (OP) associato al bruxismo (esaminare le caratteristiche neurobiologiche e fisiologiche dei muscoli mandibolari). Alcuni casi clinici e studi su piccola scala suggeriscono che alcuni farmaci legati ai sistemi dopaminergico, serotoninergico e adrenergico possono sopprimere o esacerbare il bruxismo. Inoltre, la maggior parte di questi studi farmacologici indica che varie classi di farmaci possono influenzare l'attività muscolare correlata al bruxismo, senza esercitare alcun effetto sullo OP. Pertanto, la sensibilizzazione del sistema nocicettivo del trigemino e l'effetto facilitante sui riflessi da stiramento mandibolare e sull'ipereccitabilità neurale sono fenomeni neurofisiopatogenetici correlabili al dolore nella regione craniofacciale. Tuttavia, fino ad ora, non è stata segnalata alcuna correlazione tra OP, disfunzione dei nuclei mesencefalici e facilitazione della nocicezione trigeminale, ad eccezione di uno studio clinico su un paziente affetto da cavernoma pineale, che ha evidenziato una relativa facilitazione del sistema nocicettivo trigeminale attraverso i riflessi. | ||
Come anticipato, riprenderemo lo stesso linguaggio diagnostico presentato per la paziente Mary Poppins in modo che diventi un modello assimilabile e praticabile, cercheremo di sovrapporlo al presente caso clinico denominato 'Bruxer'. Il soggetto era un uomo di 32 anni affetto da marcato bruxismo notturno e diurno e OP cronico bilaterale prevalente nelle regioni temporoparietali, con maggiore intensità e frequenza a sinistra. | Come anticipato, riprenderemo lo stesso linguaggio diagnostico presentato per la paziente Mary Poppins in modo che diventi un modello assimilabile e praticabile, cercheremo di sovrapporlo al presente caso clinico denominato 'Bruxer'. Il soggetto era un uomo di 32 anni affetto da marcato bruxismo notturno e diurno e OP cronico bilaterale prevalente nelle regioni temporoparietali, con maggiore intensità e frequenza a sinistra. | ||
=== Introduzione=== | ===Introduzione=== | ||
Come anticipato nel capitolo '[[Bruxismo]]' eviteremo di indicare questo disturbo come correlato esclusivamente dentale e cercheremo una descrizione più ampia ed essenzialmente più neurofisiologica facendo un breve excursus sui fenomeni distonici, sul 'Dolore Orofacciale' e solo successivamente considereremo il fenomeno 'bruxismo' vero e proprio. Successivamente si passerà alla presentazione del caso clinico. | Come anticipato nel capitolo '[[Bruxismo]]' eviteremo di indicare questo disturbo come correlato esclusivamente dentale e cercheremo una descrizione più ampia ed essenzialmente più neurofisiologica facendo un breve excursus sui fenomeni distonici, sul 'Dolore Orofacciale' e solo successivamente considereremo il fenomeno 'bruxismo' vero e proprio. Successivamente si passerà alla presentazione del caso clinico. | ||
| Line 234: | Line 200: | ||
File:Bruxer Jaw jerk.jpeg|'''Figure 6:''' <math>{\gamma _{2}}=</math> Strappo mandibolare rilevato elettrofisiologicamente sui masseteri destro (tracce superiori) e sinistro (tracce inferiori). La morfologia e la durata dei periodi di silenzio chiamati "soppressione esterocettiva" sembrano essere simmetriche. | File:Bruxer Jaw jerk.jpeg|'''Figure 6:''' <math>{\gamma _{2}}=</math> Strappo mandibolare rilevato elettrofisiologicamente sui masseteri destro (tracce superiori) e sinistro (tracce inferiori). La morfologia e la durata dei periodi di silenzio chiamati "soppressione esterocettiva" sembrano essere simmetriche. | ||
File:Bruxer SP2.jpg|'''Figure 7:''' <math>\gamma _3=</math> Periodo di silenzio meccanico rilevato elettrofisiologicamente sui masseteri destro (tracce sovrapposte superiori) e sinistro (tracce sovrapposte inferiori) | File:Bruxer SP2.jpg|'''Figure 7:''' <math>\gamma _3=</math> Periodo di silenzio meccanico rilevato elettrofisiologicamente sui masseteri destro (tracce sovrapposte superiori) e sinistro (tracce sovrapposte inferiori) | ||
</gallery></center> | </gallery></center> | ||
* | * | ||
====Demarcatore di coerenza <math>\tau</math>==== | ==== Demarcatore di coerenza <math>\tau</math>==== | ||
Come abbiamo descritto nel capitolo '1° Caso clinico: spasmo emimasticatorio' lo <math>\tau</math> è un peso specifico clinico rappresentativo, complesso da ricercare e sviluppare perché varia da disciplina a disciplina e per patologie, indispensabile per non far collidere le asserzioni logiche <math>\Im_o</math> e <math>\Im_n</math> nelle procedure diagnostiche e fondamentale per inizializzare la decrittazione del codice in linguaggio macchina. In sostanza consente di confermare la coerenza di una asserzione <math>\Im\cup\{\delta_1,\delta_2.....\delta_n\}</math> contro un altro <math>\Im\cup\{\gamma_1,\gamma_2.....\gamma_n\}</math> e viceversa, dando maggior peso alla gravità delle accuse e alla denuncia nel contesto opportuno. | Come abbiamo descritto nel capitolo '1° Caso clinico: spasmo emimasticatorio' lo <math>\tau</math> è un peso specifico clinico rappresentativo, complesso da ricercare e sviluppare perché varia da disciplina a disciplina e per patologie, indispensabile per non far collidere le asserzioni logiche <math>\Im_o</math> e <math>\Im_n</math> nelle procedure diagnostiche e fondamentale per inizializzare la decrittazione del codice in linguaggio macchina. In sostanza consente di confermare la coerenza di una asserzione <math>\Im\cup\{\delta_1,\delta_2.....\delta_n\}</math> contro un altro <math>\Im\cup\{\gamma_1,\gamma_2.....\gamma_n\}</math> e viceversa, dando maggior peso alla gravità delle accuse e alla denuncia nel contesto opportuno. | ||
| Line 288: | Line 254: | ||
Tuttavia, attraverso questo primo processo diagnostico abbiamo fatto progressi perché, contrariamente al processo codificato nelle discipline odontoiatriche, stiamo intraprendendo un processo neurofisiologico per decifrare il codice del linguaggio macchina del "bruxismo".<blockquote>Per non appesantire il discorso, tratteremo il secondo step diagnostico del modello Masticationpedia nel capitolo successivo intitolato '[[Codice criptato: Ipereccitabilità del sistema trigeminale]]'.</blockquote> | Tuttavia, attraverso questo primo processo diagnostico abbiamo fatto progressi perché, contrariamente al processo codificato nelle discipline odontoiatriche, stiamo intraprendendo un processo neurofisiologico per decifrare il codice del linguaggio macchina del "bruxismo".<blockquote>Per non appesantire il discorso, tratteremo il secondo step diagnostico del modello Masticationpedia nel capitolo successivo intitolato '[[Codice criptato: Ipereccitabilità del sistema trigeminale]]'.</blockquote> | ||
==Sintesi== | |||
Il bruxismo è una condizione comune che si manifesta come un disturbo del movimento, caratterizzato da serramento o digrignamento involontario dei denti, e può verificarsi sia durante la veglia che durante il sonno. Nel contesto clinico, si associa spesso a dolore orofacciale (OP), disordini temporomandibolari (TMD) e talvolta a patologie neurologiche. La difficoltà nella gestione clinica del bruxismo risiede nella sua eziologia complessa e multifattoriale, che include elementi neurologici, psicologici, dentali e neurofisiologici. | |||
== Introduzione == Come anticipato nel capitolo '[[Bruxismo]]', eviteremo di considerare il bruxismo esclusivamente come un disturbo dentale. Questa condizione richiede una visione più ampia e neurofisiologica, che includa anche i fenomeni distonici e il dolore orofacciale (OP). Nel corso di questa analisi, tratteremo non solo le cause muscolari e dentali, ma anche il coinvolgimento del sistema nervoso centrale (SNC), in particolare il sistema trigeminale. | |||
[[File | |||
.jpg|thumb|300x300px|'''Figura 1:''' Il soggetto è un uomo di 32 anni affetto da marcato bruxismo notturno e diurno e dolore orofacciale cronico bilaterale]] | |||
== Caratteristiche del Bruxismo == La distonia, che può essere associata al bruxismo, è una contrazione muscolare involontaria che può coinvolgere vari muscoli, inclusi quelli orofacciali e mandibolari. La distonia orofacciale (OFD) è una forma di distonia che interessa i muscoli facciali, orali e mandibolari, provocando movimenti involontari come il serramento eccessivo o la chiusura involontaria della mascella <ref>Thompson PD, Obeso JA, Delgado G, Gallego J, Marsden CD. 1986</ref>. Nonostante la sua eziologia sia variabile, i fattori comuni includono predisposizione genetica, traumi al SNC, farmaci, malattie neurodegenerative e disfunzioni neurotrasmettitoriali. | |||
Il bruxismo del sonno, in particolare, ha un'eziologia ancora in parte sconosciuta, ma vi sono correlazioni con disturbi del sistema neurotrasmettitoriale, come quelli legati ai sistemi dopaminergico, serotoninergico e adrenergico. Tuttavia, i farmaci che agiscono su questi sistemi sembrano influenzare solo l'attività muscolare senza impattare il dolore orofacciale associato. Il dolore cronico che i pazienti bruxisti spesso riferiscono nelle regioni temporoparietali è stato correlato a fenomeni di sensibilizzazione del sistema nocicettivo del trigemino <ref>Dubner R, Ren K. 2004</ref>. | |||
Il trattamento del bruxismo è indicato principalmente quando esso provoca dolore o danni al sistema masticatorio, come l'usura dei denti, fratture o dislocazioni articolari. Tuttavia, come evidenziato da alcuni studi, i fattori psicologici e fisiopatologici sembrano avere un ruolo molto più importante rispetto ai fattori morfologici dentali nella genesi del bruxismo <ref>Lobbezoo F. 2007</ref>. | |||
== Caso Clinico == Il paziente in esame, denominato "Bruxer," è un uomo di 32 anni che presenta bruxismo marcato, sia notturno che diurno, con dolore orofacciale bilaterale cronico, prevalentemente nelle regioni temporoparietali. Il dolore è più intenso e frequente sul lato sinistro. All'esame neurologico si è osservata una contrazione marcata dei muscoli masseteri, rigidità della mandibola, diplopia, perdita di acuità visiva nell'occhio sinistro, nistagmo con una componente rotatoria e positività del segno di Babinski bilaterale. | |||
L'analisi iniziale ha incluso indagini di tipo odontoiatrico e neurologico per differenziare la natura del disturbo. Le indagini radiologiche, come la risonanza magnetica (RM), non hanno mostrato anomalie nelle articolazioni temporomandibolari (ATM), e i tracciati assiografici condilari erano normali, come riportato in Figura 2 e Figura 3. L'elettromiografia (EMG) ha evidenziato una simmetria bilaterale nel reclutamento delle unità motorie, come visibile in Figura 4. | |||
<center><gallery widths="130" heights="200" perrow="5"> File:SC-05-0011.jpeg|'''Figura 2:''' RM dell'ATM senza segni di anomalia File:SC-05-0021.jpeg|'''Figura 3:''' Tracciato assiografico del condilo destro File:Bruxer EMG.jpeg|'''Figura 4:''' Tracciato EMG del massetere sinistro e destro </gallery></center> | |||
Il contesto odontoiatrico è stato esaminato attraverso vari parametri clinici, come i tracciati radiologici e assiografici, che sono risultati normali (<math>\delta_n=0</math> per tutti i parametri). Questo ha suggerito che, nonostante la presenza di bruxismo e dolore, il sistema dentale e muscolare masticatorio non presentava segni evidenti di patologia. | |||
== Approccio Neurologico == Il contesto neurologico ha fornito risultati più significativi. I potenziali evocati motori (MEP) delle radici trigeminali erano normali (Figura 5), ma il test del jaw jerk ha mostrato un'anomalia di ampiezza nel lato sinistro, suggerendo una possibile iperreflessia (Figura 6). Il periodo silente rilevato elettrofisiologicamente nei masseteri era simmetrico e normale (Figura 7). | |||
<center><gallery widths="250" heights="200" perrow="3"> File:Bruxer MEP.jpeg|'''Figura 5:''' Potenziali Evocati Motori del trigemino File:Bruxer Jaw jerk.jpeg|'''Figura 6:''' Strappo mandibolare (jaw jerk) con asimmetria a sinistra File:Bruxer SP2.jpg|'''Figura 7:''' Periodo silente meccanico </gallery></center> | |||
== Discussione Diagnostica == Il quadro clinico del paziente suggerisce una sovrapposizione tra aspetti odontoiatrici e neurologici. La coerenza diagnostica tra i due contesti è stata analizzata attraverso un "demarcatore di coerenza" (<math>\tau</math>), che tiene conto della gravità delle anomalie rilevate. Nel caso del paziente "Bruxer", il contesto odontoiatrico (<math>\Im_o</math>) è risultato clinicamente normale, con un valore di <math>\bar{\delta_n}=0</math>. Al contrario, il contesto neurologico (<math>\Im_n</math>) ha mostrato anomalie significative, con un valore di <math>\bar{\gamma_n}=0.33</math>, dovuto principalmente all'asimmetria del jaw jerk. | |||
Il demarcatore di coerenza ha assegnato un peso maggiore (<math>\tau_n=1</math>) al contesto neurologico, confermando una leggera pendenza diagnostica verso una componente neurologica piuttosto che odontoiatrica. Questa valutazione ha indicato che il paziente soffriva di una forma di iperreflessia trigeminale, che giustificava la persistenza del dolore orofacciale e del bruxismo cronico. | |||
== Conclusioni == Il caso clinico "Bruxer" ha evidenziato l'importanza di un approccio multidisciplinare nella diagnosi e gestione del bruxismo. Sebbene inizialmente sembrasse trattarsi di un disturbo puramente odontoiatrico, l'analisi neurologica ha rivelato una componente significativa di iperreflessia trigeminale, che potrebbe essere la causa principale del dolore e dei sintomi associati. Questo caso dimostra la necessità di integrare test neurofisiologici e odontoiatrici per una diagnosi completa e accurata del bruxismo. | |||
In futuro, ulteriori studi clinici potrebbero aiutare a chiarire le interazioni tra bruxismo e disfunzioni del sistema nervoso centrale, portando a terapie più efficaci per i pazienti che soffrono di dolore orofacciale cronico.<ref>Fernandes G, Franco AL, Gonçalves DA, Speciali JG, Bigal ME, Camparis CM. 2013</ref> | |||
* | * | ||
Latest revision as of 19:26, 20 October 2024
Se leggiamo attentamente sia le motivazioni del Research Diagnostic Criteria propenso ad eliminare le procedure cliniche di 'Stimolazione Elettrica Transcutanea' a fronte delle considerazioni di efficacia terapeutica e di supporto clinico nelle riabilitazioni del sistema masticatorio dei lavori sperimentali presentati nei capitoli precedenti non si può che rimanere 'basiti' perchè la Scienza, con tutte le sue dinamicità e rivoluzioni interne non può generare, comunque, confusione epistemologica causando a valle difficoltà ed errori diagnostici.
Per questo motivo pensiamo sia dovuto un po di chiarezza. Il primo sospettato è la 'Interdisciplinarità' intesa come la conoscenza della scienza al di fuori della propria specializzazione, infatto, in neurofisiologia il tomo muscolare non si rileva con un esame EMG di superficie. Riportiamo un esempio clinico per essere più chiari nell'asserzione.
In figura 1 viene riportato una serie di tracciati EMG di una paziente sofferente di algie orofacciali con particolare localizzazione nel massetere di sinistra ( per maggiori dettagli vedi capitoloXXXX). Il quadro elettromiografico dei masseteri è stato determinato seguendo una logica sequenza riportata nella figura 1. Come si può osservare, l’asimmetria di lato dell’attività EMG con elettrodi di superficie dei masseteri in stato di rilassamento (fig.1A) con la mandibola mantenuta in posizione di riposo era tale da richiedere una EMG ad ago del massetere sinistro. L’attività registrata con questa tecnica (fig.1B) ha mostrato una scarica con frequenza stabile di 20 Hz il che presuppose uno studio dell’unità motoria. Lo studio delle unità motorie del massetere sinistro (fig.1C), attraverso ago coassiale, ha automaticamente selezionato 26 unità motorie di cui sono state analizzate la forma, la durata, gli spikes ed i turns di ciascuna unità. I dati sono riportati nella tabella (fig.1D) Statisticamente si possono rilevare i seguenti parametri: ampiezza media di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \approxeq348\mu V} , una durata di 8.7 mS., il 23% di unità polifasiche. Questo quadro clinico rappresenta il tipico fenomeno fisiopatologico in cui il paziente riferisce dolore ma molto spesso la diagnosi rimane “difficoltà nel rilassamento muscolare”, “ dolore orofacciale atipico” o ancor meglio “fibromialgia” e di conseguenza la terapia farmacologica rimane quella sintomatica.

Figura 1a: EMG di superficie che mostra una ampia attività di scarica delle unità motorie (MUs) nel muscolo massetere di sinistra
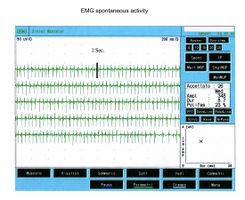
Figura 1b: Analisi della frequenza di scarica delle unità motorie masseteri (MUs)

Figura 1c: Analisi della morfologia della Unità motoria masseteri (MUs)

Figura 1d: Calcolo dei parametri di scarica delle MUs



Quello che inconfutabilmente evoca questo paragrafo da un punto di vista scientifico è che nel campo odontoiatrico, troppo concentrato nelle proprie dogmi specialistici, si interpreta l'attività EMG a riposocome tono muscolare e da qui si giunge alla eventuale diagnosi di ipertono e contestualmente trattamento con TENS. Se osserviamo attentamente la figura Ia

Lo studio di Edward Stanek IV[1] esplora il ruolo dei neuroni bilaterali proiettanti (BPN) del nucleo sopratrigeminale (SupV) nel controllo del tono basale e dell'attivazione fasica dei muscoli mandibolari, concentrandosi sui muscoli coinvolti nella chiusura della mandibola. Il documento sottolinea come i BPN possano mediare sia un'attivazione bilaterale fasica dei muscoli masseteri durante i movimenti di chiusura mandibolare, sia mantenere un livello di tono muscolare bilanciato e ridotto attraverso un'inibizione tonica.
== Introduzione == I muscoli mandibolari sono fondamentali per numerosi comportamenti orofacciali, inclusi la masticazione, la respirazione e il parlare. Tuttavia, le disfunzioni in queste attività possono portare a condizioni patologiche come il bruxismo e i disturbi dell'articolazione temporo-mandibolare[2]. Un elemento cruciale per il controllo della mandibola è la coordinazione bilaterale, che richiede un'attivazione simultanea e sincronizzata dei muscoli mandibolari su entrambi i lati. Il SupV e i nuclei reticolari parvicellulari (Rt-MoVII), situati dorsalmente rispetto al nucleo motore facciale, contengono BPN che proiettano simultaneamente ai motoneuroni mandibolari bilaterali[3]. Il SupV, in particolare, svolge un ruolo chiave nel mediare riflessi orofacciali e sensoriali, come dimostrato da studi precedenti, che ne hanno evidenziato il coinvolgimento nell'inibizione motoria dei riflessi mandibolari dolorifici[4][5].
Questo studio mira a caratterizzare i BPN di SupV e Rt-MoVII, identificandone la neurochimica, i percorsi di proiezione e il ruolo funzionale nei movimenti mandibolari, utilizzando tecniche avanzate di manipolazione genetica e optogenetica[6].
== Metodi == Per raggiungere questo obiettivo, il team di ricerca ha combinato tecniche di tracciamento virale retrogrado con il sistema split-Cre, consentendo di isolare selettivamente e manipolare i BPN[7]. L'espressione di proteine optogenetiche come Channelrhodopsin-2 (ChR2) ha permesso l'attivazione controllata dei BPN, mentre l'uso della tossina tetanica (TeNT) ha consentito di silenziarli in modo permanente, rivelando le conseguenze funzionali dell'inattività di questi neuroni sui movimenti della mandibola.
L'ibridazione in situ ha dimostrato che la popolazione di BPN è mista, contenendo neuroni eccitatori (glutammatergici) e inibitori (GABAergici e glicinergici)[8][9]. Le proiezioni di questi neuroni non si limitano ai nuclei motori trigeminali, ma coinvolgono anche altri gruppi motori orofacciali e regioni mesencefaliche implicate nelle funzioni motorie[10][11].
== Risultati == === Funzioni di SupV e Rt-MoVII === I BPN di SupV sono fondamentali per il mantenimento del tono basale del muscolo massetere, come evidenziato dal fatto che il loro silenziamento provoca un aumento significativo del tono muscolare bilaterale, suggerendo che questi neuroni esercitano un'inibizione tonica sui motoneuroni mandibolari. Questo effetto è mediato da input inibitori GABAergici e glicinergici, che regolano il livello di attività dei motoneuroni[12][13]. Tuttavia, i BPN eccitatori di SupV giocano anche un ruolo cruciale durante i movimenti fasici della mandibola, come quelli coinvolti nella masticazione, in cui l'attivazione dei BPN induce scoppi sincronizzati di attivazione muscolare bilaterale[14].
Rt-MoVII, al contrario, sembra avere un ruolo minore nella regolazione diretta dei movimenti di chiusura mandibolare. Nonostante la presenza di BPN in quest'area, il silenziamento di Rt-MoVII non ha prodotto effetti significativi sull'attivazione muscolare, suggerendo che i circuiti paralleli possono compensare per la loro funzione[15][16].
=== Manipolazione Optogenetica e Silenziamento Cronico === Gli esperimenti di attivazione optogenetica mediante ChR2 hanno rivelato che l'attivazione dei BPN eccitatori di SupV può sincronizzarsi con i movimenti naturali di chiusura della mandibola, amplificando l'attività muscolare[17]. In particolare, l'attivazione di questi neuroni durante periodi di riposo ha provocato scoppi di attivazione muscolare simili a quelli osservati durante la masticazione.
D'altra parte, il silenziamento cronico dei BPN con TeNT non ha indotto cambiamenti immediati nel tono muscolare basale, suggerendo che i meccanismi compensatori possono attenuare gli effetti dell'inibizione cronica, riducendo l'impatto del silenziamento dei BPN a lungo termine[18].
== Discussione == I risultati dello studio supportano l'idea che i BPN di SupV esercitino un duplice controllo sull'attivazione muscolare mandibolare: da un lato, essi sono responsabili di mantenere un tono muscolare bilanciato attraverso un'inibizione tonica; dall'altro, consentono attivazioni fasiche bilaterali, come quelle necessarie per la masticazione[19]. Il bilanciamento tra questi due input, eccitatorio e inibitorio, è cruciale per garantire un funzionamento normale della mandibola e prevenire disfunzioni muscolari che possono portare a condizioni patologiche come il bruxismo e altri disturbi orofacciali[20].
In sintesi, i BPN di SupV sono una popolazione mista di neuroni eccitatori e inibitori, che regolano finemente i movimenti della mandibola e l'attività dei muscoli orofacciali. L'attivazione optogenetica dimostra il loro ruolo nel generare scoppi di attivazione muscolare sincronizzati con i movimenti naturali della mandibola, mentre il silenziamento cronico e optogenetico rivela il loro contributo al mantenimento di un tono muscolare appropriato. Saranno necessari studi futuri per comprendere come queste reti neuronali siano influenzate da input sensoriali e centrali, e come i BPN contribuiscano ai disturbi orofacciali associati al sonno e ai movimenti orali alterati.
Contents

5° Clinical case: Spontaneous Electromyographic Activity
Paziente femmina di età 65 anni che riferisce principalmente Dolore Orofacciale (OP)nell'emirato sinistro della faccia ed in particolare un dolore che si irradia dai masseteri alla TMJ ed al muscolo temporale di sinistra. Dopo circa 2 anni da un episodio di perdita improvvisa di coscienza al momento in cui il proprio dentista eseguì una trocleare per paradontologia dell'emigrata inferiore sinistra. Da quel momento iniziarono dolori improvvisi di tipo untorio e poi diffusi a tutta l'emifaccia sinistra anche nella masticazione. I colleghi visto la correlazione con la masticazione ed analizzata seguendo il protocollo RDC definirono la paziente affetta da Temporomandibular Disorders (TMDs)
Giunta alla nostra attenzione seguimmo tutti i test gnatologici ( assiografia, Immagini delle ATMs, ed EMG di superficie) che non deponevano per una TMDs ma per un quadro non definito ma sostanzialmente neurologico. Il motivo di questa interpretazione fu proprio l'esecuzione della EMG di superficie che restituì i seguenti risultati. Il quadro elettromiografico dei masseteri è stato determinato seguendo una logica sequenza riportata nella figura 1. Come si può osservare, l’asimmetria di lato dell’attività EMG con elettrodi di superficie dei masseteri in stato di rilassamento (fig.1A) con la mandibola mantenuta in posizione di riposo era tale da richiedere una EMG ad ago del massetere sinistro. L’attività registrata con questa tecnica (fig.1B) ha mostrato una scarica con frequenza stabile di 20 Hz il che presuppose uno studio dell’unità motoria. Lo studio delle unità motorie del massetere sinistro (fig.1C) ha automaticamente selezionato 26 unità motorie di cui sono state analizzate la forma, la durata, gli spikes ed i turns di ciascuna unità. I dati sono riportati nella tabella (fig.1D) Statisticamente si possono rilevare i seguenti parametri: ampiezza media di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \approxeq348\mu V} , una durata di 8.7 mS., il 23% di unità polifasiche. Questo quadro clinico rappresenta il tipico fenomeno fisiopatologico in cui il paziente riferisce dolore ma molto spesso la diagnosi rimane “difficoltà nel rilassamento muscolare”, “ dolore orofacciale atipico” o ancor meglio “fibromialgia” e di conseguenza la terapia farmacologica rimane quella sintomatica. Proprio queste condizioni dovrebbero dare l’opportunità al medico di approfondire le ricerche eseguendo e conoscendo almeno in linee generali le costituenti di una analisi EMG ad ago coassiale, prima di indirizzare la paziente allo specialista neurologo.
Figura 1a: EMG di superficie che mostra una ampia attività di scarica delle unità motorie (MUs) nel muscolo massetere di sinistra
Figura 1b: Analisi della frequenza di scarica delle unità motorie masseteri (MUs)
Figura 1c: Analisi della morfologia della Unità motoria masseteri (MUs)
Figura 1d: Calcolo dei parametri di scarica delle MUs
Steps EMG ad ago
L’esame EMG dei muscoli scheletrici consiste di quattro step:
- Attività di inserzione quando inserito l’elettrodo ad ago nel muscolo
- Attività spontanea quando il muscolo si trova in condizioni di riposo
- Potenziali di unità motoria evocate da isolate scariche motorie durante una moderata contrazione volontaria
- Reclutamento o pattern interferenziale durante progressivo livello di contrazione
Attività di inserzione
In un soggetto l’attività di inserzione appare come spikes ad alta frequnenza positivi e negativi in un unico gruppo e sono in genere la rappresentazione di un danno delle fibre muscolari o di una stimolazione meccanica dovuta alla penetrazione dell’ago nel muscolo. Nel nostro paziente questa attività si presentò con una durata di 80 mS e fu riferibile ad una quadro di normalità. Da notare, inoltre, il fenomeno dell’attività di placca. Se un elettrodo ad ago viene mantenuto stazionario in un punto del muscolo, i muscoli normali a riposo non mostrano assolutamente nessuna attività elettrica eccetto che nella regione della placca neuromuscolare. Queste consistono di due componenti: bassa ampiezza ( nell’ordine di 10-50 μV) e minima durata (1-2 msec) che all’altoparlante EMG assomigliano al suono delle conchiglie di mare sull’orecchio. Nel nostro caso (fig.1A) l’assenza totale di attività di placca nel massetere destro può essere spiegata dalla registrazione eseguita con elettrodi di superficie che abbattono parzialmente l’energia del segnale ma l’attività registrata sul massetere sinistro, sempre con elettrodi di superficie, ha un ampiezza di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \approxeq100\mu V} . Per lo stesso ragionamento questa attività non deve essere considerata come attività di placca in quanto, come si può osservare in fig. 1B, registrazione del massetere sinistro eseguita con elettrodo coassiale, l’ampiezza è di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \approxeq 400\mu V} . A volte gli spikes di potenziali di placca sono indistinguibili nella forma d’onda dai potenziali di fibrillazione i quali anche mostrano una iniziale negatività quando registrati vicino alla placca. Un altro elemento curioso è la somiglianza del modello di scarica tra le scariche dei fusi neuromuscolari e dei potenziali di placca, tanto è vero che alcuni autori [21] ipotizzarono che questi potenziali potessero originare dalle fibre intrafusali muscolari. Rimane ancora aperta la discussione ed il significato elettrofisiologico da dare all’attività elettrica osservabile in fig. 1B.
Attività spontanea:

Nelle prime 2 settimane dopo la denervazione la sensibilità di una fibra muscolare all’acetilcolina (ACh) aumenta fina a 100 volte. Questo fenomeno conosciuto come “ipersensibilità da denervazione” può spiegare le scariche spontanee delle fibre muscolari denervate in risposta a minimi quanti di ACh. Il fatto che l’infusione di curaro blocca i recettori della placca neuromuscolare ma non abolisce la scarica spontanea[22], che la denervazione del muscolo di rana può determinare una aumentata sensibilità alla ACh ma non generare attività spontanea [23]. Questi studi hanno suggerito una ipotesi alternative quella dei cambiamenti lenti dei potenziali di membrana di origine metabolica che possono periodicamente raggiungere un livello critico ed evocare spikes propagati.[24] I fenomeni tipici dell’attività spontanee comprendono, comunque, potenziali di fibrillazione, onde a punta positiva, potenziali di fascicolazione, scariche miochimiche e scariche ripetitive complesse. Senza entrare in argomenti troppo specialistici e considerando la registrazioni elettrofisiologiche del caso clinico è sufficiente trattare le onde a punta positive, la fibrillazione e la fascicolazione. Per onde punta positive si intende scariche a denti di sega che scaricano spontaneamente e continuamente. Questo tipo di attività si trova nelle muscoli denervati ma anche in una varietà di condizioni miogeniche. In figura 2 si può osservare un tipico tracciato di attività spontanea di onde a punta positiva che confrontate con il caso clinico in esame (fig.1B) sono chiaramente diverse. Per fibrillazione si intende, invece, potenziali di durata da Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 1-5} Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle msec} e ampiezza di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \approxeq 20-200\mu V} con forme d’onda bifasiche o trifasiche e positività iniziale. Potenziali di fibrillazione triggerati da oscillazioni spontanee nel potenziale di membrana tipicamente scaricano a frequenze di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 1-30 Hz} con una media di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 13Hz} . Questo fenomeno rappresenta l’attività spontanea di una o più fibre muscolari ed è patognomonico di denervazione anche se può apparire in muscoli sani. La presenza di scariche riproducibili in almeno due differenti aree del muscolo usualmente suggerisce un disordine del motoneurone secondario che include patologie delle cellule delle corna anteriori, radicolopatie, plessopatie, mono e polineuropatie assonali oltre che in certe miopatie.

In figura 3 possiamo osservare un tipico tracciato di attività spontanea da denervazione e confrontarlo con il tracciato in figura 1C in cui si possono notare delle diversità elettrofisiologiche. L’attività spontanea di fibrillazione ha un ampiezza di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \approxeq 200\mu V} , la frequenza risulta essere Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 13Hz} con andamento random mentre nel caso clinico riportato (fig.1C) l’ampiezza era di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \approxeq 400\mu V} e la frequenza più alta (Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \approxeq 13Hz} ) ma particolarmente stabile quasi a significare un pacemaker centrale.
Per evitare confusione terminologica e clinica tra fibrillazione e fascicolazione Danny-Brown e Pennybacker[25] proposero il termine di fascicolazione per descrivere la spontanea contrazione (twitch) di unità motorie. Le fascicolazione, perciò, rappresentano la scarica spontanea di un gruppo di fibre muscolari riferibili all’intera o parziale parte dell’unità motoria. Isolate scariche di una unità motoria con complesse bursts di ripetitive scariche causano movimenti vermicolari della pelle chiamati miochimie.[26]

Scariche ripetitive della stessa unità motoria occorrono in bursts a intervalli regolari di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 0.1-10sec} con Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 2-10} spikes che scaricano a Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 30-40 Hz} in ciascun burst. I potenziali di fascicolazione sono tipicamente associati con patologie delle cellule delle corna anteriori ma sono anche osservate in radicolopatie, neuropatie da intrappolamento, e la sindrome di fascicolazione dolorosa muscolare. In figura 4 si può osservare un chiaro esempio di fascicolazioni del muscolo orbicolare dell’occhio che confrontato con i tracciati del caso clinico (fig.1B e C) mostra una totale diversità morfologica e di rappresentazione temporale. Questa diversità rafforzerebbe l’esclusione di una patologia da denervazione.
Potenziali di Unità Motoria
Una unità motoria può essere definita dall’ampiezza, il rise time, dalla durata e dalle fasi come verrà meglio descritto nel capitolo 'Elettromiografia'. L’ampiezza registrata varia ampiamente con la posizione della punta dell’elettrodo relativa alla sorgente di corrente ionica scaricata, per cui un operatore esperto seleziona un potenziale di unità motoria con una rise time di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \approxeq 500\mu sec} per essere certi della vicinanza con la sorgente. L’ampiezza nel range della normalità va da centinaia di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \mu V} ad alcuni Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle m V} e la durata da Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 5-10 msec} . Per i muscoli facciali, in particolare, ci si riferisce ai valori riportati da Buchthal [27] il cui range è Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 4,2-7,5 msec} per una età massima di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 75} anni. I potenziali di unità motoria bifasici o trifasici sono presenti anche in muscoli normali con una media di Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 5-15%} di unità con Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 4} o più fasi.

Il numero di unità polifasiche aumentano sia nelle miopatie, neuropatie o nelle patologie del motoneurone. La polifasia indica, dunque, una dispersione temporale dei potenziali di fibra muscolari dentro una unità motoria. In alcune anormalità chiamate doppiette o triplette una unità motoria spara due o tre volte ad una intervallo di tempo molto corto e sono rappresentative di un disturbo metabolico associato ad ipereccitabilità del pool motoneurale. In figura 5 possiamo osservare un tipico tracciato a minima attività volontaria di MUAP polifasico ed una doppietta che rappresenta uno stato di patologia del motoneurone. Confrontando questa registrazione di unità motoria patologica con alcune della fig.1C, ed in particolare la 5,7,13 e 23 con i valori di ampiezza rispettivi Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 678\mu V;419\mu V3;686\mu V e 530\mu V} durata Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 8.2, 4.2,6.2 msec } e e 8.4 Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle 8.4 msec } possiamo affermare che l’attività registrata sul massetere di sinistra, del caso clinico in questione, non ha nessuna caratteristica elettrofisiologica sovrapponibile ad un quadro di danno del II motoneurone.
Sintesi
=== 5° Clinical case: Spontaneous Electromyographic Activity === Il 5° caso clinico riguarda una paziente femmina di 65 anni che lamenta dolore orofacciale (OP) nell'emirato sinistro della faccia, descritto come un dolore irradiato dai masseteri fino all'articolazione temporomandibolare (TMJ) e al muscolo temporale sinistro. Il dolore si è manifestato circa due anni dopo un episodio di perdita improvvisa di coscienza verificatosi durante una procedura paradontologica presso il suo dentista, che ha eseguito una trocleare sull'emiarcata inferiore sinistra. Da quel momento, la paziente ha iniziato a soffrire di dolori improvvisi e diffusi su tutta l'emifaccia sinistra, che si acuiscono durante la masticazione.
I colleghi, notando la correlazione con la masticazione e seguendo il protocollo Research Diagnostic Criteria (RDC), diagnosticarono alla paziente un disturbo temporomandibolare (TMDs) [28].
Alla nostra attenzione, abbiamo eseguito tutti i test gnatologici (assiografia, immagini delle articolazioni temporomandibolari, e elettromiografia di superficie) che non indicavano chiaramente la presenza di un TMDs, ma piuttosto un quadro neurologico. In particolare, l'elettromiografia (EMG) di superficie ha mostrato risultati anomali. Il tracciato elettromiografico dei masseteri è stato esaminato seguendo una logica precisa riportata in figura 1.
Come si osserva in figura 1a, l'EMG di superficie del massetere sinistro in condizioni di rilassamento con la mandibola in posizione di riposo ha evidenziato un'attività asimmetrica rispetto al lato destro. Questa asimmetria ha reso necessario eseguire un'EMG ad ago sul massetere sinistro per un'analisi più approfondita. L'elettromiografia ad ago (EMG) ha rivelato una scarica con una frequenza stabile di 20 Hz (figura 1b), suggerendo la necessità di uno studio dell'unità motoria.
L'analisi delle unità motorie del massetere sinistro (figura 1c) ha selezionato automaticamente 26 unità motorie, di cui sono state analizzate la forma, la durata, gli spike e i turns. I dati sono riassunti in figura 1d e mostrano un'ampiezza media di circa 348 μV, una durata di 8.7 ms, e il 23% di unità polifasiche.
Questo quadro clinico riflette un fenomeno fisiopatologico tipico in cui la paziente riferisce dolore, ma la diagnosi rimane spesso vaga, classificata come "difficoltà nel rilassamento muscolare," "dolore orofacciale atipico" o "fibromialgia." Di conseguenza, la terapia è generalmente sintomatica. Tuttavia, tali condizioni dovrebbero spingere il medico ad approfondire le indagini, eseguendo un'analisi EMG ad ago prima di indirizzare la paziente a uno specialista neurologo.
Figura 1d: Calcolo dei parametri di scarica delle MUs
==== Steps EMG ad ago ==== L'esame EMG dei muscoli scheletrici consiste in quattro fasi fondamentali:
Attività di inserzione al momento dell'inserimento dell'elettrodo nel muscolo. Attività spontanea quando il muscolo è in condizioni di riposo. Potenziali di unità motoria evocati da scariche isolate durante una moderata contrazione volontaria. Reclutamento o pattern interferenziale durante un progressivo livello di contrazione. ===== Attività di inserzione ===== In un soggetto normale, l'attività di inserzione si presenta come spike ad alta frequenza, sia positivi che negativi, e rappresenta una risposta a stimolazioni meccaniche dovute all'inserimento dell'ago nel muscolo. Nel caso della nostra paziente, questa attività si è manifestata con una durata di 80 ms ed è stata considerata normale.
Da notare che nei muscoli normali a riposo, non c'è alcuna attività elettrica significativa ad eccezione della regione della placca neuromuscolare, che può manifestare spike di bassa ampiezza (<10-50 μV) e di breve durata (1-2 ms) [29]. Nel nostro caso clinico, l'assenza di attività di placca sul massetere destro può essere attribuita alla registrazione con elettrodi di superficie, che possono attenuare il segnale [30]. Tuttavia, l'attività sul massetere sinistro registrata con elettrodi di superficie ha mostrato un'ampiezza di circa 100 μV, suggerendo un'origine diversa dall'attività di placca, come evidenziato in figura 1b.
===== Attività spontanea ===== [[File a punta+.jpeg|thumb|Figura 2: Attività spontanea con scariche a punta positive in un muscolo denervato]] L'attività spontanea nei muscoli denervati è comunemente osservata nelle prime settimane dopo la denervazione e può essere spiegata da un aumento della sensibilità all'acetilcolina (ACh). Questo fenomeno, noto come "ipersensibilità da denervazione," può causare scariche spontanee delle fibre muscolari in risposta a stimoli minimi di ACh[31].
Nel nostro caso, tuttavia, l'attività spontanea osservata nel massetere sinistro della paziente non presentava le caratteristiche tipiche della denervazione, come le onde a punta positiva o la fibrillazione, come illustrato in figura 2. Questi risultati hanno suggerito che l'attività spontanea registrata non fosse correlata a una patologia del motoneurone inferiore.
===== Potenziali di unità motoria ===== Le unità motorie vengono definite in base all'ampiezza, durata, rise time, e numero di fasi. Un operatore esperto può selezionare potenziali di unità motoria con un rise time di circa 500 μsec per garantire la vicinanza alla sorgente. Nei muscoli facciali, l'ampiezza normale varia tra alcuni centinaia di μV fino a pochi mV, con una durata media tra 5 e 10 ms [32]. In condizioni normali, si osserva una certa presenza di potenziali polifasici (potenziali con quattro o più fasi), ma il loro numero tende ad aumentare in caso di neuropatie o miopatie [33].
La figura 5 mostra un tracciato di attività polifasica, in cui è visibile la presenza di potenziali polifasici e doppiette. Confrontando questi risultati con le registrazioni delle unità motorie della paziente (figura 1c), possiamo escludere la presenza di una patologia del motoneurone inferiore nel caso clinico in esame.
===== Fascicolazioni e scariche ripetitive ===== [[File .jpeg|thumb|Figura 4: Fascicolazioni del muscolo orbicolaris oculi sinistro.]] Le fascicolazioni sono contrazioni spontanee di unità motorie e sono comunemente osservate in patologie delle cellule delle corna anteriori, radicolopatie, neuropatie da intrappolamento, e nella sindrome delle fascicolazioni dolorose[34]. Le scariche ripetitive della stessa unità motoria avvengono in burst regolari e sono rappresentative di disturbi del motoneurone. Come si vede in figura 4, le fascicolazioni nel muscolo orbicolare dell'occhio sinistro mostrano una morfologia differente rispetto ai tracciati EMG del massetere sinistro della paziente, confermando l'assenza di patologie da denervazione.
In conclusione, il caso clinico ha dimostrato come l'elettromiografia ad ago sia uno strumento essenziale per identificare anomalie neuromuscolari. Sebbene inizialmente il caso fosse stato diagnosticato come un disturbo temporomandibolare, l'analisi elettromiografica ha rivelato un quadro clinico più complesso, suggerendo l'importanza di una diagnosi neurologica approfondita in pazienti con dolore orofacciale atipico.
Bruxer

Pochi studi hanno tentato di caratterizzare il dolore orofacciale (OP) associato al bruxismo (esaminare le caratteristiche neurobiologiche e fisiologiche dei muscoli mandibolari). Alcuni casi clinici e studi su piccola scala suggeriscono che alcuni farmaci legati ai sistemi dopaminergico, serotoninergico e adrenergico possono sopprimere o esacerbare il bruxismo. Inoltre, la maggior parte di questi studi farmacologici indica che varie classi di farmaci possono influenzare l'attività muscolare correlata al bruxismo, senza esercitare alcun effetto sullo OP. Pertanto, la sensibilizzazione del sistema nocicettivo del trigemino e l'effetto facilitante sui riflessi da stiramento mandibolare e sull'ipereccitabilità neurale sono fenomeni neurofisiopatogenetici correlabili al dolore nella regione craniofacciale. Tuttavia, fino ad ora, non è stata segnalata alcuna correlazione tra OP, disfunzione dei nuclei mesencefalici e facilitazione della nocicezione trigeminale, ad eccezione di uno studio clinico su un paziente affetto da cavernoma pineale, che ha evidenziato una relativa facilitazione del sistema nocicettivo trigeminale attraverso i riflessi.
Come anticipato, riprenderemo lo stesso linguaggio diagnostico presentato per la paziente Mary Poppins in modo che diventi un modello assimilabile e praticabile, cercheremo di sovrapporlo al presente caso clinico denominato 'Bruxer'. Il soggetto era un uomo di 32 anni affetto da marcato bruxismo notturno e diurno e OP cronico bilaterale prevalente nelle regioni temporoparietali, con maggiore intensità e frequenza a sinistra.
Introduzione
Come anticipato nel capitolo 'Bruxismo' eviteremo di indicare questo disturbo come correlato esclusivamente dentale e cercheremo una descrizione più ampia ed essenzialmente più neurofisiologica facendo un breve excursus sui fenomeni distonici, sul 'Dolore Orofacciale' e solo successivamente considereremo il fenomeno 'bruxismo' vero e proprio. Successivamente si passerà alla presentazione del caso clinico.

La distonia è una contrazione muscolare involontaria, ripetitiva, prolungata (tonica) o spasmodica (rapida o clonica). Lo spettro delle distonie può coinvolgere varie regioni del corpo. Di interesse per i chirurghi orali e maxillofacciali sono le distonie cranio-cervicali, in particolare la distonia orofacciale (OFD). L'OFD è una contrazione involontaria e prolungata dei muscoli periorbitali, facciali, oromandibolari, faringei, laringei o cervicali.[35] L'OFD può coinvolgere i muscoli masticatori, facciali inferiori e della lingua, che possono causare trisma, bruxismo, apertura o chiusura involontaria della mascella e movimento involontario della lingua
L'eziologia dell'OFD è varia e comprende predisposizione genetica, lesioni al sistema nervoso centrale (SNC), traumi periferici, farmaci, stati metabolici o tossici e malattie neurodegenerative. Tuttavia, nella maggior parte dei pazienti, non è possibile identificare alcuna causa specifica. È stata trovata un'associazione tra disturbi temporomandibolari dolorosi (TMD), emicrania, cefalea di tipo tensivo e bruxismo notturno, sebbene l'associazione fosse significativa solo per l'emicrania cronica. L'associazione tra TMD dolorosi e bruxismo del sonno ha aumentato significativamente il rischio di emicrania cronica, seguita da emicrania episodica e cefalea di tipo tensivo episodica.[36]
Il bruxismo è il disturbo del movimento orale più frequente e può verificarsi nei soggetti durante la veglia e durante il sonno. È probabile che entrambe le forme abbiano eziologie diverse e la loro diagnosi e trattamento richiedono approcci diversi. Il trattamento è indicato quando il bruxismo provoca dolore al sistema masticatorio o porta a danni come l'usura dei denti o fratture di denti, restauri o persino di impianti. Una revisione mirata sull'eziologia del bruxismo[37] ha concluso che esiste un ruolo limitato per i fattori morfologici nell'eziologia del bruxismo, mentre i fattori psicologici (ad esempio lo stress) e i fattori fisiopatologici (ad esempio i disturbi nei sistemi di neurotrasmettitori centrali) sono maggiormente coinvolti.
Il dolore orofacciale (OP), compreso il dolore da TMD, esercita un effetto modulatore sui riflessi di stiramento mandibolare.[38] Studi elettrofisiologici hanno dimostrato che il dolore indotto sperimentalmente da iniezioni di soluzione salina ipertonica al 5% nel muscolo massetere provoca un aumento dell'ampiezza da picco a picco dello scatto mandibolare. Questo effetto facilitatore sembra essere correlato ad una maggiore sensibilità del sistema fusimotorio, che allo stesso tempo provoca rigidità muscolare.[39] Inoltre, numerosi studi su animali sul dolore muscolare indotto sperimentalmente hanno dimostrato che l'attivazione dei nocicettori muscolari influenza notevolmente le proprietà propriocettive dei fusi muscolari attraverso una via neurale centrale[40] e che il lavaggio della sostanza algogena locale provoca un ritorno alla normalità riflessi tendinei.
Tuttavia, pochi studi hanno tentato di caratterizzare il dolore associato al bruxismo (cioè, di esaminare le caratteristiche neurobiologiche e fisiologiche dei muscoli mandibolari). Alcuni casi clinici e studi su piccola scala suggeriscono che alcuni farmaci legati ai sistemi dopaminergico, serotoninergico e adrenergico possono sopprimere o esacerbare il bruxismo. Inoltre, la maggior parte di questi studi farmacologici indica che varie classi di farmaci possono influenzare l'attività muscolare correlata al bruxismo, senza esercitare alcun effetto sull'OP.[41]
Pertanto, la sensibilizzazione del sistema nocicettivo del trigemino e l'effetto facilitante sui riflessi da stiramento mandibolare e sull'ipereccitabilità del SNC sono fenomeni neurofisiopatogenetici correlabili al dolore nella regione craniofacciale. Tuttavia, fino ad ora, non è stata segnalata alcuna correlazione tra OP, disfunzione dei nuclei mesencefalici e facilitazione della nocicezione trigeminale, ad eccezione di uno studio clinico su un paziente affetto da cavernoma pontino, che ha evidenziato una relativa facilitazione del sistema nocicettivo trigeminale attraverso il riflesso delle palpebre.[42]
Caso Clinico
Come anticipato riprenderemo lo stesso linguaggio diagnostico presentato per la paziente Mary Poppins in modo che diventi un modello assimilabile e praticabile, e cercheremo di sovrapporlo al presente caso clinico denominato 'Bruxer'.
Il soggetto era un uomo di 32 anni affetto da marcato bruxismo notturno e diurno e OP cronico bilaterale prevalente nelle regioni temporoparietali, con maggiore intensità e frequenza a sinistra. L'esame neurologico ha mostrato una contrazione dei muscoli masseteri con pronunciata rigidità della mandibola, diplopia e perdita dell'acuità visiva nell'occhio sinistro, nistagmo dello sguardo sinistro con una componente rotatoria, papille con bordi sfocati e positività bilaterale di Babynski e riflessi tendinei policinetici in tutti quattro arti.
a quanto esposto nei precedenti capitoli dalla 'Introduzione' ai capitoli 'Logica del linguaggio medico' e all'ultimo capitolo 'Bruxismo', oltre alla complessità delle argomentazioni e alla vaghezza del linguaggio verbale, potremmo trovare ci troviamo di fronte ad una situazione clinica in cui sembra dominare uno dei contesti considerati.
(sembra ma....)
A differenza del paziente con 'Spasmo Emimasticatorio', il caso clinico del nostro povero 'Bruxer' mostra un fenomeno di sovrapposizione di proposizioni, asserzioni e frasi logiche nel contesto odontoiatrico e neurologico e apparentemente nessuno dei due ottiene un'assoluta e chiara compatibilità e coerenza . Questo ha delle ripercussioni nella clinica in cui tutti gli attori coinvolti (i medici legali) hanno ragione e contestualmente torto, rendendo la conclusione diagnostica inadeguata e pericolosa, ma vediamo passo dopo passo il processo nel suo complesso.
Significato dei contesti
In ambito odontoiatrico avremo le seguenti frasi ed affermazioni alle quali diamo un valore numerico per facilitare il trattamento, ovvero Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \delta_n=[0|1]} dove Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \delta_n=0} indica 'normale' e Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \delta_n=1} anomalia e quindi positività del referto:
Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \delta_1} Referto RM negativo dell'ATM nella Figura 2,Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \delta_1=0\longrightarrow} Normalità, negatività del referto
Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \delta_2} Referto assiografico negativo per tracciato condilare destro in Figura 3,Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \delta_2=0\longrightarrow} Normalità, negatività del referto
Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \delta_3} Referto assiografico negativo per tracciato condilare sinistro in Figura 4,Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \delta_3=0\longrightarrow} Normalità, negatività del referto
Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \delta_4} Diagramma di interferenza EMG simmetrico nella Figura 5,Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \delta_4 =0\longrightarrow} Normalità, negatività del referto
Figura 2: Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \delta_1=0\longrightarrow} immagini RM dell'articolazione temporo-mandibolare (lato destro mostrato solo per semplificazione) non mostrano segni di deragliamento meniscale e/o infiammazione. Rapporto risultato negativo.




(ed è proprio qui che i contesti entrano in conflitto o meglio i risultati potrebbero non essere così decisivi)
Nel contesto neurologico avremo, perciò, le seguenti frasi ed asserzioni a cui diamo un valore numerico per facilitare la trattazione e cioè Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \gamma_n=[0|1]} dove lo Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \gamma_n=0} indica 'normalità' e Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \gamma_n=1} 'anormalità e dunque positività del referto:
Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \gamma_1=} Presenza e simmetria dei Potenziali Evocati Motori delle Radici trigeminali in Figure 5, Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \gamma_0=0\longrightarrow} Normalità, negatività del referto
Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \gamma_2=} Presenza del jaw jerk con relativa asimmetria di ampiezza in Figure 6 Failed to parse (MathML with SVG or PNG fallback (recommended for modern browsers and accessibility tools): Invalid response ("Math extension cannot connect to Restbase.") from server "https://wikimedia.org/api/rest_v1/":): {\displaystyle \gamma_2=1\longrightarrow} Anormalità, negatività del referto* ( lo * è stato inserito per annotare una ambiguità del referto che andremo a descrivere dettagliatamente nella trattazione clinica)
Periodo silente elettrico e contestuale simmetria Figure 7, Normalità, negatività del referto


Figure 5: Potenziali Evocati Motori delle radici del trigemino





Demarcatore di coerenza
Come abbiamo descritto nel capitolo '1° Caso clinico: spasmo emimasticatorio' lo è un peso specifico clinico rappresentativo, complesso da ricercare e sviluppare perché varia da disciplina a disciplina e per patologie, indispensabile per non far collidere le asserzioni logiche e nelle procedure diagnostiche e fondamentale per inizializzare la decrittazione del codice in linguaggio macchina. In sostanza consente di confermare la coerenza di una asserzione contro un altro e viceversa, dando maggior peso alla gravità delle accuse e alla denuncia nel contesto opportuno.




Il peso del demarcazione , quindi, dà maggior significato alle asserzioni più gravi nel contesto clinico da cui derivano e quindi al di là della maggiore o minore positività delle asserzioni o che comunque sono sempre verificate e rispettate, queste devono essere validato in base alla gravità clinica intrinseca moltiplicando la media delle asserzioni e per uno dove indica 'bassa gravità' mentre 'alta gravità'.




![{\displaystyle \tau =[0|1]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/fbdc534cec0dcf1f070a551e40611eb83e8aca25)


Riassumendo nel nostro caso 'Bruxer' abbiamo quindi:

dove
media del valore delle affermazioni cliniche nel contesto odontoiatrico e quindi


media del valore delle affermazioni cliniche nel contesto neurologico e quindi


segnalazione di bassa gravità del contesto dentale

segnalazione di elevata gravità del contesto neurologico

dove il demarcatore di coerenza '' definirà il percorso diagnostico come segue

Come si vede nel nostro caso clinico 'Bruxer' abbiamo una leggerissima pendenza diagnostica verso il contesto neurologico che ci permette però di intravedere più una componente neurologica che dentale.

Lavata via la miriade di dati normativi riportati positivamente, che generano conflitto tra contesti, grazie al demarcatore di coerenza abbiamo un quadro molto più chiaro e lineare su cui approfondire l'analisi della funzionalità del Sistema Nervoso Centrale rispetto nel nostro caso clinico 'Bruxer' appare alquanto incuriosito dal basso peso diagnostico derivato dalle asserzioni neurologiche .

Questa cifra media deriva principalmente da un'ipotetica anomalia dell'ampiezza dello scatto della mandibola etichettata con un asterisco (*). Ne parleremo nella sezione dedicata a questo riflesso trigeminale.
Di conseguenza ci si può concentrare sull'intercettazione dei test necessari per decifrare il codice del linguaggio macchina che il SNC invia all'esterno convertito in linguaggio verbale che a prima vista sembrerebbe riguardare una sorta di iperreflessia dei riflessi tendinei. e in particolare lo scatto della mascella..[43][44][45] Per confermare questa ipotetica intuizione è necessario un brainstorming del tipo 'Cognitive Neural Network' abbreviato in 'RNC' presentato per la diagnosi del caso della nostra 'Mary Poppins' nel capitolo 'Codice cifrato: trasmissione efaptica'.
Tuttavia, attraverso questo primo processo diagnostico abbiamo fatto progressi perché, contrariamente al processo codificato nelle discipline odontoiatriche, stiamo intraprendendo un processo neurofisiologico per decifrare il codice del linguaggio macchina del "bruxismo".
Per non appesantire il discorso, tratteremo il secondo step diagnostico del modello Masticationpedia nel capitolo successivo intitolato 'Codice criptato: Ipereccitabilità del sistema trigeminale'.
Sintesi
Il bruxismo è una condizione comune che si manifesta come un disturbo del movimento, caratterizzato da serramento o digrignamento involontario dei denti, e può verificarsi sia durante la veglia che durante il sonno. Nel contesto clinico, si associa spesso a dolore orofacciale (OP), disordini temporomandibolari (TMD) e talvolta a patologie neurologiche. La difficoltà nella gestione clinica del bruxismo risiede nella sua eziologia complessa e multifattoriale, che include elementi neurologici, psicologici, dentali e neurofisiologici.
== Introduzione == Come anticipato nel capitolo 'Bruxismo', eviteremo di considerare il bruxismo esclusivamente come un disturbo dentale. Questa condizione richiede una visione più ampia e neurofisiologica, che includa anche i fenomeni distonici e il dolore orofacciale (OP). Nel corso di questa analisi, tratteremo non solo le cause muscolari e dentali, ma anche il coinvolgimento del sistema nervoso centrale (SNC), in particolare il sistema trigeminale.
[[File .jpg|thumb|300x300px|Figura 1: Il soggetto è un uomo di 32 anni affetto da marcato bruxismo notturno e diurno e dolore orofacciale cronico bilaterale]]
== Caratteristiche del Bruxismo == La distonia, che può essere associata al bruxismo, è una contrazione muscolare involontaria che può coinvolgere vari muscoli, inclusi quelli orofacciali e mandibolari. La distonia orofacciale (OFD) è una forma di distonia che interessa i muscoli facciali, orali e mandibolari, provocando movimenti involontari come il serramento eccessivo o la chiusura involontaria della mascella [46]. Nonostante la sua eziologia sia variabile, i fattori comuni includono predisposizione genetica, traumi al SNC, farmaci, malattie neurodegenerative e disfunzioni neurotrasmettitoriali.
Il bruxismo del sonno, in particolare, ha un'eziologia ancora in parte sconosciuta, ma vi sono correlazioni con disturbi del sistema neurotrasmettitoriale, come quelli legati ai sistemi dopaminergico, serotoninergico e adrenergico. Tuttavia, i farmaci che agiscono su questi sistemi sembrano influenzare solo l'attività muscolare senza impattare il dolore orofacciale associato. Il dolore cronico che i pazienti bruxisti spesso riferiscono nelle regioni temporoparietali è stato correlato a fenomeni di sensibilizzazione del sistema nocicettivo del trigemino [47].
Il trattamento del bruxismo è indicato principalmente quando esso provoca dolore o danni al sistema masticatorio, come l'usura dei denti, fratture o dislocazioni articolari. Tuttavia, come evidenziato da alcuni studi, i fattori psicologici e fisiopatologici sembrano avere un ruolo molto più importante rispetto ai fattori morfologici dentali nella genesi del bruxismo [48].
== Caso Clinico == Il paziente in esame, denominato "Bruxer," è un uomo di 32 anni che presenta bruxismo marcato, sia notturno che diurno, con dolore orofacciale bilaterale cronico, prevalentemente nelle regioni temporoparietali. Il dolore è più intenso e frequente sul lato sinistro. All'esame neurologico si è osservata una contrazione marcata dei muscoli masseteri, rigidità della mandibola, diplopia, perdita di acuità visiva nell'occhio sinistro, nistagmo con una componente rotatoria e positività del segno di Babinski bilaterale.
L'analisi iniziale ha incluso indagini di tipo odontoiatrico e neurologico per differenziare la natura del disturbo. Le indagini radiologiche, come la risonanza magnetica (RM), non hanno mostrato anomalie nelle articolazioni temporomandibolari (ATM), e i tracciati assiografici condilari erano normali, come riportato in Figura 2 e Figura 3. L'elettromiografia (EMG) ha evidenziato una simmetria bilaterale nel reclutamento delle unità motorie, come visibile in Figura 4.
Figura 4: Tracciato EMG del massetere sinistro e destro
Il contesto odontoiatrico è stato esaminato attraverso vari parametri clinici, come i tracciati radiologici e assiografici, che sono risultati normali ( per tutti i parametri). Questo ha suggerito che, nonostante la presenza di bruxismo e dolore, il sistema dentale e muscolare masticatorio non presentava segni evidenti di patologia.

== Approccio Neurologico == Il contesto neurologico ha fornito risultati più significativi. I potenziali evocati motori (MEP) delle radici trigeminali erano normali (Figura 5), ma il test del jaw jerk ha mostrato un'anomalia di ampiezza nel lato sinistro, suggerendo una possibile iperreflessia (Figura 6). Il periodo silente rilevato elettrofisiologicamente nei masseteri era simmetrico e normale (Figura 7).
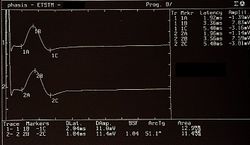
Figura 7: Periodo silente meccanico
== Discussione Diagnostica == Il quadro clinico del paziente suggerisce una sovrapposizione tra aspetti odontoiatrici e neurologici. La coerenza diagnostica tra i due contesti è stata analizzata attraverso un "demarcatore di coerenza" (), che tiene conto della gravità delle anomalie rilevate. Nel caso del paziente "Bruxer", il contesto odontoiatrico () è risultato clinicamente normale, con un valore di . Al contrario, il contesto neurologico () ha mostrato anomalie significative, con un valore di , dovuto principalmente all'asimmetria del jaw jerk.


Il demarcatore di coerenza ha assegnato un peso maggiore () al contesto neurologico, confermando una leggera pendenza diagnostica verso una componente neurologica piuttosto che odontoiatrica. Questa valutazione ha indicato che il paziente soffriva di una forma di iperreflessia trigeminale, che giustificava la persistenza del dolore orofacciale e del bruxismo cronico.
== Conclusioni == Il caso clinico "Bruxer" ha evidenziato l'importanza di un approccio multidisciplinare nella diagnosi e gestione del bruxismo. Sebbene inizialmente sembrasse trattarsi di un disturbo puramente odontoiatrico, l'analisi neurologica ha rivelato una componente significativa di iperreflessia trigeminale, che potrebbe essere la causa principale del dolore e dei sintomi associati. Questo caso dimostra la necessità di integrare test neurofisiologici e odontoiatrici per una diagnosi completa e accurata del bruxismo.
In futuro, ulteriori studi clinici potrebbero aiutare a chiarire le interazioni tra bruxismo e disfunzioni del sistema nervoso centrale, portando a terapie più efficaci per i pazienti che soffrono di dolore orofacciale cronico.[49]
- ↑ Edward Stanek 4th 1, Erica Rodriguez 1, Shengli Zhao 1, Bao-Xia Han 1, Fan Wang 2. Supratrigeminal Bilaterally Projecting Neurons Maintain Basal Tone and Enable Bilateral Phasic Activation of Jaw-Closing Muscles. J Neurosci. 2016 Jul 20;36(29):7663-75. doi: 10.1523/JNEUROSCI.0839-16.2016.
- ↑ Balasubramaniam R, Ram S. 2008
- ↑ Li et al., 1993
- ↑ Kamogawa et al., 1994
- ↑ Minkels et al., 1995
- ↑ Stanek et al., 2014
- ↑ Wang et al., 2012
- ↑ Travers et al., 2005
- ↑ Paik et al., 2009
- ↑ Fay e Norgren, 1997
- ↑ Kolta et al., 2000
- ↑ Brooks e Peever, 2008
- ↑ Gemba-Nishimura et al., 2010
- ↑ Inoue et al., 1994
- ↑ Yamamoto et al., 1989
- ↑ Inoue et al., 1992
- ↑ Madisen et al., 2012
- ↑ Yamamoto et al., 2003
- ↑ Travers et al., 2005
- ↑ Richter e Löscher, 1998
- ↑ Partanen JV, Nousiainen U. : End-plate spikes in the electromyography are fusimotor unit potentials. Neurology 1983; 33:039-1043,
- ↑ Axelsson J, Thesleff S.: A study of super-sensitivity of denervated mammalian skeletal muscles. J Physiol (Lond) 1957:149;178-193
- ↑ Miledi R.: The acetylcholine sensitivity of frog musclefibres after complete or partial denervation. J Physiol (Lond) 1960:151;1-23
- ↑ Thesleff S: Fibrillation in denervated mammalian skeletal muscle. In Cukp WL e Ochoa J (eds): Abnormal Nerves and Muscle as Impulse Genertors. Oxford University Press. Oxford 1982, pp 678-694
- ↑ Danny-Brown D, Pennybacker JB.: Fibrillation and fasciculation in voluntary muscle. Brain 1938; 61: 311-332
- ↑ Sindermann F,Conrad B, Jacobi HM, Prochazka VJ.: Unusual properties of repetitive fasciculation Elctroencephalogr Clin Neurophysiol 1973; 35: 173-179
- ↑ Buchtal F: An introduction to electromyography Scandinavian University Books. Copenhagen 1957
- ↑ Balasubramaniam R, Ram S. 2008
- ↑ Axelsson J, Thesleff S. 1957
- ↑ Miledi R. 1960
- ↑ Thesleff S. 1982
- ↑ Buchthal F. 1957
- ↑ Partanen JV, Nousiainen U. 1983
- ↑ Sindermann F, Conrad B. 1973
- ↑ Thompson PD, Obeso JA, Delgado G, Gallego J, Marsden CD. Focal dystonia of the jaw and differential diagnosis of unilateral jaw and masticatory spasm. J Neurol Neurosurg Psychiatry. 1986;49:651–656. doi: 10.1136/jnnp.49.6.651. [PMC free article][PubMed] [CrossRef] [Google Scholar][Ref list]
- ↑ Fernandes G, Franco AL, Gonçalves DA, Speciali JG, Bigal ME, Camparis CM. Temporomandibular disorders, sleep bruxism, and primary headaches are mutually associated. J Orofac Pain. 2013;27(1):14–20. [PubMed] [Google Scholar] [Ref list]
- ↑ Lobbezoo F. Taking up challenges at the interface of wear and tear. J Dent Res. 2007;86(2):101–103. doi: 10.1177/154405910708600201.[PubMed] [CrossRef] [Google Scholar][Ref list]
- ↑ Dubner R, Ren K. Brainstem mechanisms of persistent pain following injury. J Orofac Pain. 2004;18(4):299–305. [PubMed] [Google Scholar] [Ref list]
- ↑ Wang K, Svensson P, Arendt-Nielsen L. Modulation of exteroceptive suppression periods in human jaw-closing muscles by local and remote experimental muscle pain. Pain. 1999;82(3):253–262. doi: 10.1016/S0304-3959(99)00058-5.[PubMed] [CrossRef] [Google Scholar][Ref list]
- ↑ Ro JY, Capra NF. Modulation of jaw muscle spindle afferent activity following intramuscular injections with hypertonic saline. Pain. 2001;92(1–2):117–127.[PubMed] [Google Scholar] [Ref list]
- ↑ Winocur E, Gavish A, Voikovitch M, Emodi-Perlman A, Eli I. Drugs and bruxism: a critical review. J Orofac Pain. 2003;17(2):99–111. [PubMed] [Google Scholar] [Ref list]
- ↑ Katsarava Z, Egelhof T, Kaube H, Diener HC, Limmroth V. Symptomatic migraine and sensitization of trigeminal nociception associated with contralateral pontine cavernoma. Pain. 2003;105(1–2):381–384.[PubMed] [Google Scholar] [Ref list]
- ↑ S Watanabe , H Mochizuki, I Nakashima, Y Itoyama. A case of primary Sjögren's syndrome with CNS disease mimicking chronic progressive multiple sclerosis.Rinsho Shinkeigaku. 1998 Jul;38(7):658-62.
- ↑ Ibrahim M Norlinah, Kailash P Bhatia, Karen Ostergaard, Robin Howard, Gennarina Arabia, Niall P Quinn. Primary lateral sclerosis mimicking atypical parkinsonism. Mov Disord. 2007 Oct.31;22(14):2057-62. doi: 10.1002/mds.21645.
- ↑ M Yoshida, N Murakami, Y Hashizume, A Takahashi. A clinicopathological study on 13 cases of motor neuron disease with dementia.Rinsho Shinkeigaku.1992 Nov;32(11):1193-202.
- ↑ Thompson PD, Obeso JA, Delgado G, Gallego J, Marsden CD. 1986
- ↑ Dubner R, Ren K. 2004
- ↑ Lobbezoo F. 2007
- ↑ Fernandes G, Franco AL, Gonçalves DA, Speciali JG, Bigal ME, Camparis CM. 2013